The protocol described above covers the preparation of Drosophila pupae for long-term live imaging and the procedures for the analysis of cell orientation and growth dynamics of the abdominal epidermis. By applying this methodology it is possible to generate high-resolution movies of the developing pupae for periods of up to 48 h without significant photobleaching or phototoxicity. Snapshots depicting the abdominal epidermis (e.g., histoblasts and LECs) at different time points and from pupae oriented at different angles are shown in Figure 4. The subsequent analysis of these movies allows identification and quantification of the dynamics of local and global changes in the main geometrical and shape parameters modulated during the expansion of histoblasts and the replacement of LECs. These analyses can be performed in different scenarios and specific mutant backgrounds. They can also be employed for clones in which gene expression is altered, leading to autonomous loss or gain of function conditions. This would allow the exploration of nonautonomous responses in surrounding cells, facilitating the identification of cross talk or cell communication mechanisms (Figure 5). This approach has been recently employed in the identification of the Fat/Dachsous/Four jointed pathway as a key element directing and orienting cells alignment during the deploying of the planar polar pattern of the abdominal epidermis of the adult9.

Figure 1: Dissecting and mounting pupae for live imaging. (A) From left to right: dorsal view of a prepupa at 0 h APF (left) and of a pupa at 14 h APF before (middle) and after (right) the removal of the opaque pupal case. The opercular region is indicated (white arrowheads). (B) The essential toolkit for dissection is shown. From left to right: glass slide, double-sided sticky tape, a pair of forceps, and a glass-bottom dish. (C) Staged pupae are immobilized on double-sided sticky tape on glass slides dorsal side up. The pupal case of each pupa is opened from the opercular region with surgical forceps. (D−E) The peeling of the pupal case is gradual. The case is torn and folded to the pupal sides from the head to the abdomen. (F−H) The pupa is gently lifted from the ventral side with the tips of the forceps (G) and transferred to a glass-bottom dish over a minimal drop of halocarbon oil. (I) The pupa is then appropriately oriented (dorsolaterally, top; dorsally, bottom). Multiple pupae can be mounted simultaneously by repeating the steps shown in panels C−I. (J) Image showing the outline of the cells of the dorsal histoblast nest of the AIII segment expressing the junctional marker Atpα::GFP. The pupa was oriented dorsolaterally and imaged at 14 h APF. Scale bar = 22 µm. (K) Image equivalent to (J) but from a dorsal point of view. The dynamics of tissue development can be visualized for several hours. See also Figure 4. Please click here to view a larger version of this figure.
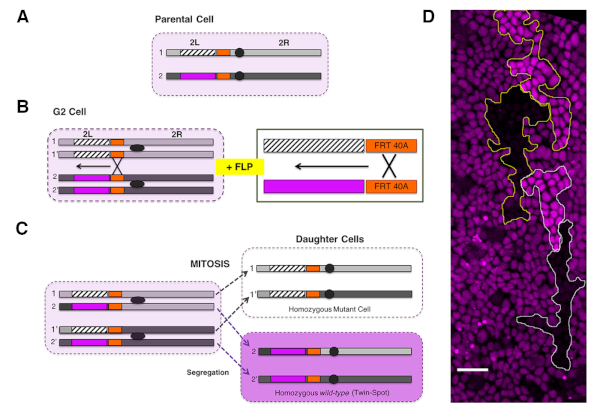
Figure 2: Generation of genetic mosaics via FLP/FRT system. (A) A parental heterozygous cell (pale magenta) carries a recessive mutant allele (dashed rectangle) on one chromosome arm and a gene encoding for a fluorescent marker (magenta) in the other chromosome arm [the left arm of the chromosome 2 (2L) in this example]. FRT sites (orange rectangles) are engineered in both arms at the same chromosomal positions proximal to the centromere (grey ovals). (B) The activation of FLP recombinase by heat shock in cells paused in G2 leads to the recombination between the FRTs of sister chromatids 1-1' and 2-2'. As a result, the region distal to the FRT sites (FRT40A on 2L) is exchanged. A magnified view is shown on the right panel. (C) Upon the polar segregation of the rearranged regions (1-1' and 2-2') during mitosis, two genetically different sister cells are generated. One daughter is homozygous for the recessive allele and for the entire chromosome arm distal to the site of recombination. This cell lacks the gene that encodes for the fluorescent marker, negatively marked in white. The other daughter cell will be homozygous for the wild type arm, giving rise to a twin-spot and expressing two copies of the genes encoding for the fluorescent marker (dark magenta). For simplicity, segregations of rearranged regions to opposite poles of the mitotic cell (i.e., 1−2 and 1'−2') are not shown. These give rise to cells phenotypically indistinguishable from the parental cell (heterozygous background, pale magenta). (D) Image showing clones in the A compartment generated via FLP/FRT somatic recombination at the third instar larvae stage and visualized at 26 h APF. Clones of cells are marked by the absence of RFP.nls (black) and homozygous expression of RFP.nls (bright magenta, twin spots) in an otherwise heterozygous RFP.nls background (dim magenta). Equivalent events can be achieved for FRT sites located in other chromosomal locations (2R, 3L, 3R, and X). See also Figure 5. Please click here to view a larger version of this figure.
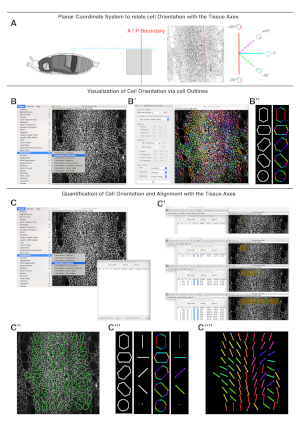
Figure 3: Illustration of cell orientation measurements. To extract information on cell orientations, first a planar coordinate system to relate cell orientation with tissue axis is set (A). Second, cell orientations are extracted from cell outlines (B). Last, cell orientations and alignments with the tissue axis are quantified (C). (A) On the left, a diagram of a lateral view of a pupa oriented according to the planar coordinate system. The red solid and the cyan dashed lines define the Cartesian plane representing the anteroposterior (A/P) boundary and the dorsal midline respectively. In the middle, an inverted image showing a dorsolateral view of the expanding histoblasts at 26 h APF outlined by the junctional marker Atpα::GFP (input image). The position of the A/P boundary is highlighted with a red line. On the right, an axial compass showing the color code applied to describe cell edge orientations (polygons) or averaged cell orientations (bars). (B) Display of the Plugins/OrientationJ menu and the OrientationJ Distribution window. (B') Display of the OrientationJ Distribution window showing the settings of the parameters employed to obtain the cell edge orientation map on the right. (B'') Illustration of the color-coded cell outlines on idealized cells. A circle does not show any preferred color. (C) Display of the OrientationJ Measure window. (C') Sequential screenshots of the OrientationJ Measure window showing how to measure local orientation and coherency in consecutive ROIs of uniform weight. The local orientation angle and coherency for each region are displayed as ellipsoids. (C'') Representation of the final results of the orientation measurement. Ellipsoids visually display the orientation (i.e., angle of the ellipsoid longest axis with respect to the A/P boundary) and coherency (i.e., ratio between the longest and the shorter axis of the ellipsoid). The numerical values of both parameters are saved in a spreadsheet for further analyses. (C''') Illustration of the color-coded cell outlines and averaged cell orientation (bars) on idealized cells. A circle does not show any preferred orientation. (C'''') Representation of the preferred local orientation of each ROI (locally averaged orientation map). The colors highlight the orientation of each region. Anterior is to the left. Scale bar = 22 μm. Please click here to view a larger version of this figure.
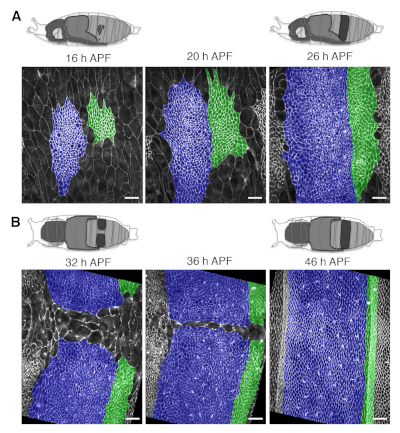
Figure 4: Long-term live imaging of growing abdominal epithelia. (A) Representative snapshots from long-term imaging movies from early to late phases of histoblast expansion. Top: Schematic views of a pupa oriented for dorsolateral imaging at 16 h and 26 h APF. The territory occupied by the nests visible from the dorsolateral side of the pupa is highlighted in dark grey at 16 and 26 h APF. Bottom: Images showing cell outlines from both histoblasts and LECs labeled by the ubiquitous expression of the junctional marker Atpα::GFP. The A/P boundary lies in between the two highlighted compartments (Fake colored cells in blue = anterior compartment; green = posterior compartments). (B) Representative snapshots from long-term imaging movies from early to late phases of nests confluence. Top: Schematic view of a pupa oriented for dorsal imaging at 32 h and 48 h APF. Bottom: Cell outlines (labeled and colored as in A). Note that the Atpα::GFP marker allows delineation of the shape of individual epithelial cells over time with high resolution. Scale bar = 22 μm. Please click here to view a larger version of this figure.
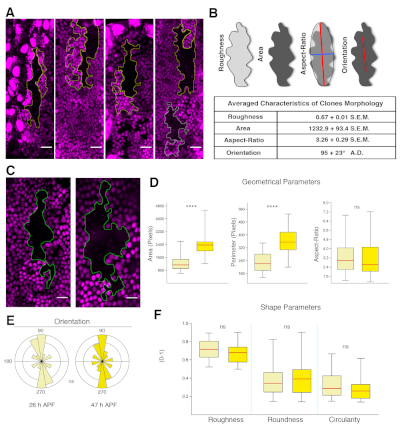
Figure 5: Tissue properties extracted from clonal analysis. (A) Examples of wild type clones in the A compartment marked by the absence of RFP.nls (black) and their twin spots (bright magenta) at 26 h APF. The clones elongate along the segmental boundaries. Twin clones arrange in parallel or in tandem. Scale bar = 22 μm. (B) Top: Images illustrating the parameters quantified from the clone outlines. Bottom: Summary table reporting the average values for the indicated parameters for wild type animals (n = 29). (C) Morphology of a wild type clone at 26 h (left) and 47 h (right) APF. The clone shows complex border morphology at both stages. (D) Box and whisker plots for geometrical parameters at 26 h (light yellow) and 47 h APF (dark yellow). The averaged area and perimeter increase significantly in this time window. (E) Polar plots representing the clones orientation (bin size 18°, abundance proportional to the area). Orientation is sustained during expansion and remodeling. (F) Box and whisker plots for shape parameters at 26 h (light yellow) and 47 h APF (dark yellow). Roughness (solidity), roundness, and circularity barely change. The median values are shown with a red horizontal line and whiskers extend to the minimum and maximum values of the distribution. Statistics was performed with K-SM test or W-tests (p < 0.0001****, p > 0.05 not significant). Anterior is to the left, dorsal is up. Scale bar = 16 μm. Genotype is hsflp1.22;FRT40A/FRT40A Ubi.RFP.nls. Please click here to view a larger version of this figure.
