All the animal procedures were performed in accordance with guidelines from the Danish Animal Experimentation Inspectorate and approved by the local ethical committee for Experimental Animals.
1. Tissue processing
NOTE: Following transcardial perfusion and post fixation in 4% paraformaldehyde (PFA) diluted in 0.2 M phosphate buffer overnight at 4 °C, brains are transferred to 30% sucrose in phosphate buffered saline (PBS) for three days at 4 °C. Thereafter, the brains are frozen on powdered dry ice and stored at -80 °C until sectioning.
- Separate the right and left hemispheres along the midline of the brain using a scalpel and then choose one or the other hemisphere randomly in the first brain, thereafter shift systematically between right and left hemisphere (Figure 1A). Mount the sampled hemisphere onto a specimen disc with the long axis perpendicular to the disc using a mounting medium.
- Place numbered multidish containers for pre-cooling in the cryostat.
- Mount the specimen disc into the cryostat and cut the entirety of the hippocampus (Bregma 1.80 to - 7.0416) into 80 µm thick sections in the coronal plane (Figure 1B).
- Place all sampled sections consecutively in the cooled multidish containers, to ensure that the sections are kept in cutting-order.
- Cover the sections completely with cryoprotectant and hold the containers at -20 °C until further use.
2. Immunostaining
NOTE: To keep track of the individual sections throughout the staining procedure, place the sampled sections in 25-well staining nets. Use every 5th section, giving a mean final number of 12 (8-16) sections per hippocampus for immunostaining and subsequent cell counting.
- Prior to subsampling every nth section with a random start between section one and section n (Figure 1C) transfer the containers from -20 °C to room temperature (RT).
- Use e.g. a paintbrush, to transfer the sections in numerical order to the 25-well staining nets placed in petri dishes filled with PBS.
NOTE: From this point, keep the sections in the staining nets which are placed in matching glass dishes placed on an orbital shaker (section 2.3 to 2.9). - Transfer the staining nets to the matching glass dishes and wash the sections twice for 10 min in PBS at RT before incubation in 3% hydrogen peroxide (H2O2) diluted in distilled water (dH2O) for 20 min.
- Transfer the sections to three different solutions of hydrochloric acid (HCl) (1 M at 0 °C for 10 min, 2 M at RT for 10 min, and 2 M at 37 °C for 20 min) before neutralization in 0.1 M sodium tetraborate at RT for 20 min.
- After 3 × 10 min incubations in 1% Triton X-100 diluted in PBS (washing buffer), transfer the sections to washing buffer containing 10% fetal bovine serum (blocking solution) for 1 h at RT.
- Incubate the sections for 48 h at 4 °C in mouse-anti-BrdU diluted 1:100 in blocking solution.
- Wash the sections 3 × 10 min in washing buffer, and incubate for 48 h in horse radish peroxidase diluted 1:10 in washing buffer.
- Transfer the sections to PBS for 5 × 10 min, and then for 7 min in 0.01% diaminobenzidine (DAB) diluted in PBS before they are transferred to a similar DAB solution containing 0.02% H2O2 for 10 min at RT.
- Complete a series of washes (2 × 10 min in PBS and 2 × 10 min in phosphate buffer without added sodium chloride) and mount the sections in numerical order on microscope slides.
- Dry the sections for approximately 30 min before counterstaining with cresyl violet.
- Place the microscope slides in a slide rack.
- Rehydrate the sections for 10 min in slide staining dishes containing dH2O, and then transfer the slides to 0.02% cresyl violet in dH2O for 15 min.
- Repeat step 2.12 before the sections are dehydrated three times in ethanol (96% for 5 min and 99% for 2 × 2 min).
- Place the sections in xylene for 2 × 15 min and cover-slip the slides using a rapid drying medium for mounting.
- Finally, leave the cover-slipped slides for approximately 24 h before they can be used for microscopy.
3. Estimation of the total number of BrdU-labelled neurons using the optical fractionator
NOTE: Conduct a pilot study including a few animals to determine the optimal sampling parameters, such as the number of sections to be analyzed and the number of optical disectors within the sampled sections. This pilot also provides a preliminary CV (standard deviation/mean) and the possibility to adjust the CE (see section 3.9.3) to obtain a satisfactory precision of the estimates determined by the investigator (for more details see below). Likewise perform a z-distribution analysis to address the following points: shrinkage of the tissue, quality of the staining throughout the section and distribution of the cells in the z-axis14.
- Check the thickness of the tissue in the sections to confirm that they are suitable for the use of the optical fractionator, e.g. is thick enough for the chosen disector height including guard zones (see below). NOTE: The tissue thickness is measures in several places within the region of interest.
- Place the slides on the motorized stage of the microscope and turn on the preferred stereological software.
- Delineate the area of interest using a low-magnification objective (2X or 4X) before changing to a 100X oil-immersion objective (see Figure 2A).
- Using a specific point of a counting frame (e.g. at or adjacent to a corner), locate the top of the section by moving along the focal plane until some feature of the section appears in focus. Register this z-position as 0 (see Figure 2B).
- Move the focal plane down through the tissue until the same specific point of the counting frame is at the last z-level of tissue in focus and mark this position. The local tissue thickness is defined from 0 to this endpoint and can be read on the z-axis. Register the tissue thickness (see Figure 2B).
- Document full penetration of the stain and distribution of the cells in the full thickness of the section.
NOTE: The distribution of BrdU-labelled neurons is used as a guide for determining the required guard zones. Although a uniform distribution of cells is critical, it is acceptable to observe fewer cells near the top and bottom of the section, indicating the effect of lost caps (for further details see Dorph-Petersen et al., 2009).- Sample BrdU-labelled neurons and register their z-position together with the local section thickness measured in the selected corner of each counting frame.
- Plot the number of BrdU-positive neurons as a function of their z-position.
- Fix the height of the disector and guard zones based on the mean section thickness and the distribution of cells (see 3.1 and 3.2).
NOTE: The height of the disector should be less than the section thickness to avoid artifacts close to the surfaces. This risk is circumvented by including guard zones to the top and bottom of the section. - Determine the step length between the disectors.
- Delineate the region of interest on all the sections to be analyzed and register the areas.
- Sum these areas and then divide by the number of disectors to be placed within this area.
NOTE: Based on previous experience a good starting point entails 75 probes. This will provide an approximation of the area and the corresponding sampling positions (Astep) (for further details see West, 2012). - Take the square root of Astep, which will provide the x-y-step size.
- Determine the size of the counting frame.
- Set the size of the counting frame to an arbitrary unit and perform a pilot sampling of BrdU-positive neurons using the parameters obtained in section 3.3 and 3.4.
NOTE: The size has to be determined empirically by trial and error, but it is recommended that it is adjusted to enable sampling of 2-3 cells per disector8.
- Set the size of the counting frame to an arbitrary unit and perform a pilot sampling of BrdU-positive neurons using the parameters obtained in section 3.3 and 3.4.
- Following this final step of the pilot study begin cell sampling.
NOTE: The obtained sampling parameters should result in counts of approximately 150-200 cells in each animal which is sufficient to obtain an efficient stereological design8. - Count the BrdU-positive neurons using a 100× oil-immersion objective and a final magnification of 2000-3000×. In each optical disector, identify any BrdU-labelled neurons that are clearly recognized by the feature of interest (in the present study, the criterion is when the leading edge of the BrdU-labelled neuron comes into focus for the first time), are located inside the counting frame or touch the inclusion lines (Figure 2B). Do not count the BrdU-positive neurons that, when clearly recognized by the feature of interest inside the disector height, touch the exclusion lines. It is critically important to follow scrupulously these counting rules throughout the entire stereological quantification.
NOTE: As BrdU incorporates into all dividing cells, morphology is used to distinguish between neurons and other types of cells. Neuron characteristics are a clearly defined nucleus with a neutral cytoplasm and a dark nucleolus. BrdU-positive cells are recognized as newly formed neurons based on their relatively larger size compared to glial cells. It should be mentioned that in studies of short-term proliferation (e.g. < 24 h) double-staining using immature neuronal markers may be a prerequisite17. - Estimate the total number of BrdU-positive neurons in each brain by multiplying the total number of cells counted (ΣQ–) with the reciprocal sampling fractions:
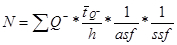
for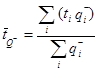
where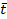 Q- is the number-weighted mean section thickness, h is the height of the disector, asf is the area sampling fraction, ssf is the section sampling fraction and ti is the section thickness in the ith counting frame with a cell count of
Q- is the number-weighted mean section thickness, h is the height of the disector, asf is the area sampling fraction, ssf is the section sampling fraction and ti is the section thickness in the ith counting frame with a cell count of 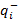 in the disector14,18.
in the disector14,18. - Calculate the coefficient of error (CE)8 which is the sampling variance of BrdU-positive neurons in systematic uniformly random sampling (SURS).
- First, calculate the noise, which is equal to the total number of BrdU-positive neurons counted:
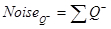
- Next, calculate the variance in SURS:
NOTE: We have judged the area and cell counts in the hippocampus to change smoothly from section to section and consequently used "smoothness class" m = 1 (1/240)8,19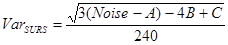
where
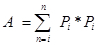
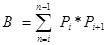
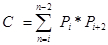
Pi is the number of points counted for the object in the ith section and n is the number of sections4,20. - Finally, calculate the CE of the estimate:
NOTE: In general, optimal precision is typically achieved when the CE value is less than half of the observed CV, as OCV2 = ICV2 + CE2, where OCV is the observed CV and ICV is the inherent CV4.
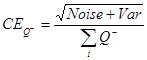
- First, calculate the noise, which is equal to the total number of BrdU-positive neurons counted:
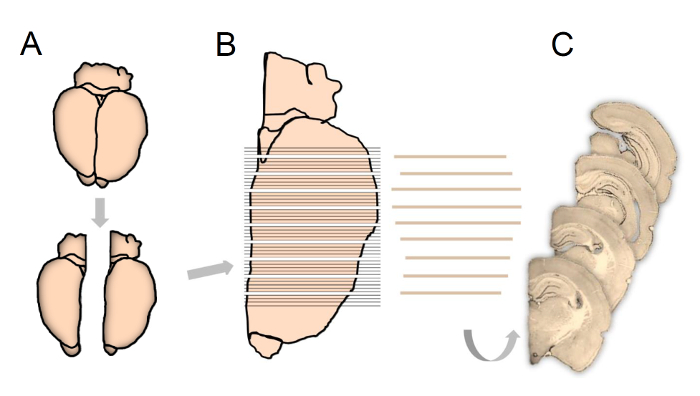
Figure 1: Schematic illustration showing tissue processing according to the fractionator principles used in the present study.
Following experimentation, the hemispheres are separated and right or left hemisphere chosen systematically and randomly (A). The complete brain extending over the entire hippocampus is cut into 80-µm-thick coronal sections (B), whereupon every 5th section is sub-sampled, starting randomly between section 1 to 5, for final immunostaining and stereological quantification (C). Please click here to view a larger version of this figure.
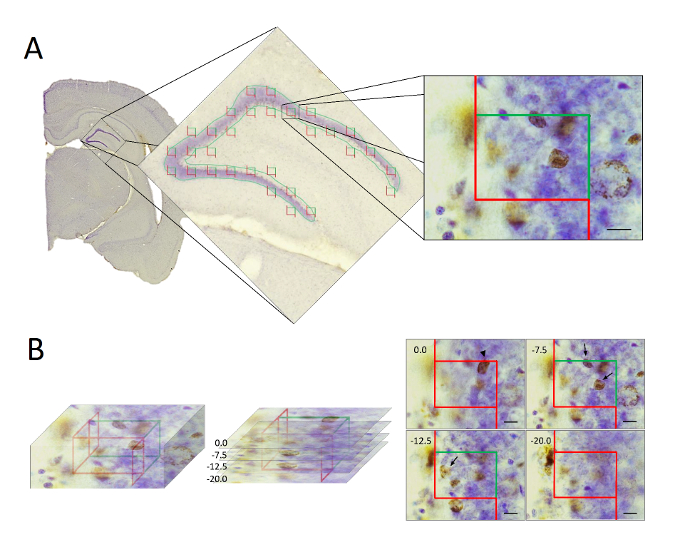
Figure 2: Illustration showing the stereological sampling procedure using optical disectors.
(A) Following histological processing the hippocampal GCL/SGZ was delineate and optical disectors applied uniformly and randomly over the region. (B) In the optical disectors (left), the BrdU-positive neurons were counted only if their feature of interest were clearly recognized within the disector height, and located inside the counting frame or touching the inclusion line (right). Neurons touching the exclusion line were not counted. Scale bars = 10 µm. Please click here to view a larger version of this figure.
Sampling parameters
By analyzing the z-distribution in a pilot study (Figure 3), we found a uniform distribution of BrdU-positive neurons in the disector height. Shrinkage of the sections was measured; final mean thickness was 26.4 μm (19.0-32.0 μm) in sections originally cut at 80 μm. Based on the tissue thickness and distribution of BrdU labelled neurons the disector height was set to 10 μm, and the guard zones at the top and bottom of the section was set to 5-μm and 4-17 μm respectively. Depending of the density of the cells, the area of the counting frames was 1414 or 5210 μm2. The step length was 220 μm in both the x and y directions and cell counting was performed in an average of 12 (range 8-16) sections per animal. These sampling parameters resulted in a mean of 133 (range 33-372) BrdU-positive cells counted in 173 (107-204) disectors in each brain hemisphere.
As an example, we obtained the following CV and CE values for BrdU-positive neuronal estimations in ECS treated rats: 24 hours: CV = 0.59, CE = 0.11; 3 months: CV = 0.34, CE = 0.12; 6 months: CV = 0.43, CE = 0.09; 12 months: CV = 0.22, CE = 0.09.
We quantified the total number of newly-formed neurons and their long-term survival in the rat hippocampus following ECS using the optical fractionator in conjunction with a BrdU staining technique10,11. The final results showed a basal neurogenesis in the four groups of control animals, which did not significantly differ as a function of survival time (Figure 3). Furthermore, ECS induced a significant increase in the total number of BrdU-positive neurons at 24 hours (259%, p<0.0001), 3 (229%, p<0.001), 6 (152%, p<0.05) and 12 months (162%, p<0.05) following experimentation, as compared to controls animals.
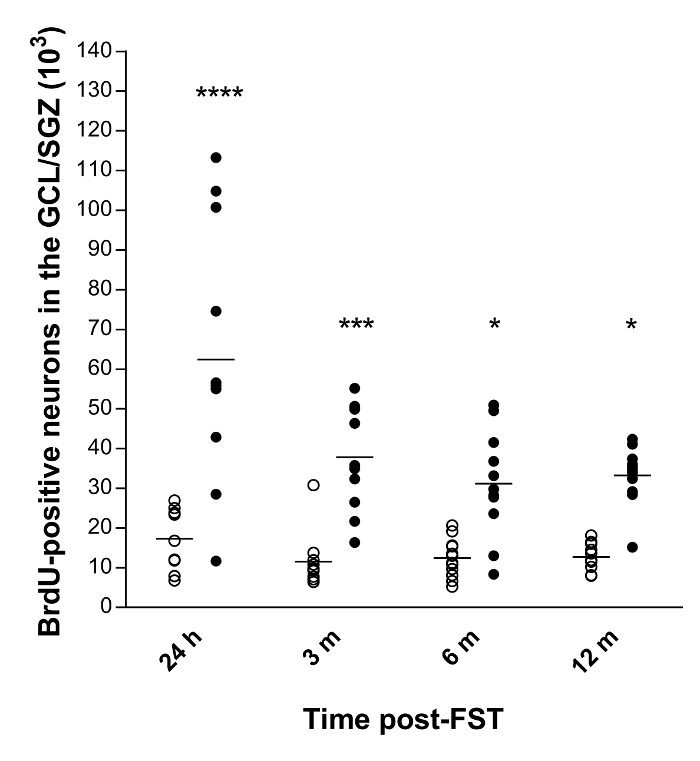
Figure 3: Quantitative data of the total number of BrdU-positive neurons in the GCL/SGZ of the rat hippocampus10,11.
Horizontal bars represent the mean values. Abbreviations: GCL, granule cell layer; SGZ, sub-granular zone; h, hours; m, months. ****p<0.0001, ***p<0.001 and *p<0.05 versus control (n=11-12 per group). Please click here to view a larger version of this figure.
