In this method paper plasmid DNA consisting of green fluorescent protein (GFP) expression cassettes was targeted into the developing chicken basilar papilla (BP) with optimized parameters of 12 V and 4 pulses with 100 msec pulse duration and intervals of 200 msec, yielding a ~50% embryo survival rate and efficiency of plasmid-DNA targeting into the BP. Fluorescent imaging of native GFP expression in developing embryos showed this method of electroporation preferentially targets the anterior-ventral aspect of the otocyst, that gives rise to the BP (Figure 1 E-E'; white arrow) and GFP expression can be followed over a time course of up to approximately ~4 days (Figure 1 E-H). RNA in situ hybridization (ISH) was used to determine whether the electroporation successfully targeted the auditory portion of the developing inner ear. To determine whether GFP was taken up by auditory sensory progenitors, ISH experiments were performed on adjacent BP sections. Within the developing BP sensory progenitors were identified by their expression of Sox2 (Figure 1 K and N) and Lunatic fringe (Lfng) (Figure 1 J). These experiments demonstrate that sensory progenitors throughout the developing BP, as shown for the base (Figure 1 I; black bracket) and apex (Figure 1 L; black bracket), were successfully targeted and express GFP after 68 hr of electroporation (Figure 1 I and L; black brackets). In addition, GFP was expressed outside the sensory domain in non-sensory epithelial cells (Figure 1 I and L; red arrows) and in the auditory ganglion (Figure 1I and L; ag, yellow arrows) marked by Neurod1 expression (Figure 1M). In a typical experiment GFP expression in targeted cells persisted for up to ~4 days (Figure 1 H); this allows the investigator to analyze the spatial and temporal pattern of sensory progenitor cell-cycle withdrawal and differentiation in the developing BP. To determine the proliferative behavior of sensory progenitors, the thymidine analog EdU was added at the time of electroporation at E4. Four days later, EdU incorporation in the developing BP was analyzed in tissue sections using "click" chemistry. Immuno-labeling for the HC-specific protein MyosinVIIa (Myo7a) was used to identify differentiating HCs in the developing BP. In the apical portion of the BP Myo7a+ HCs were frequently EdU+ (Figure 1 P-P'; P'', white arrows), whereas in the base only few Myo7a+ HCs had incorporated EdU (Figure 1 O-O'; O'', white arrows), suggesting that at the time of EdU addition at E4, sensory progenitors in the base are already largely post mitotic. These results confirm previous studies of opposing gradients of cell-cycle exit and HC differentiation in the chick BP 6,7.
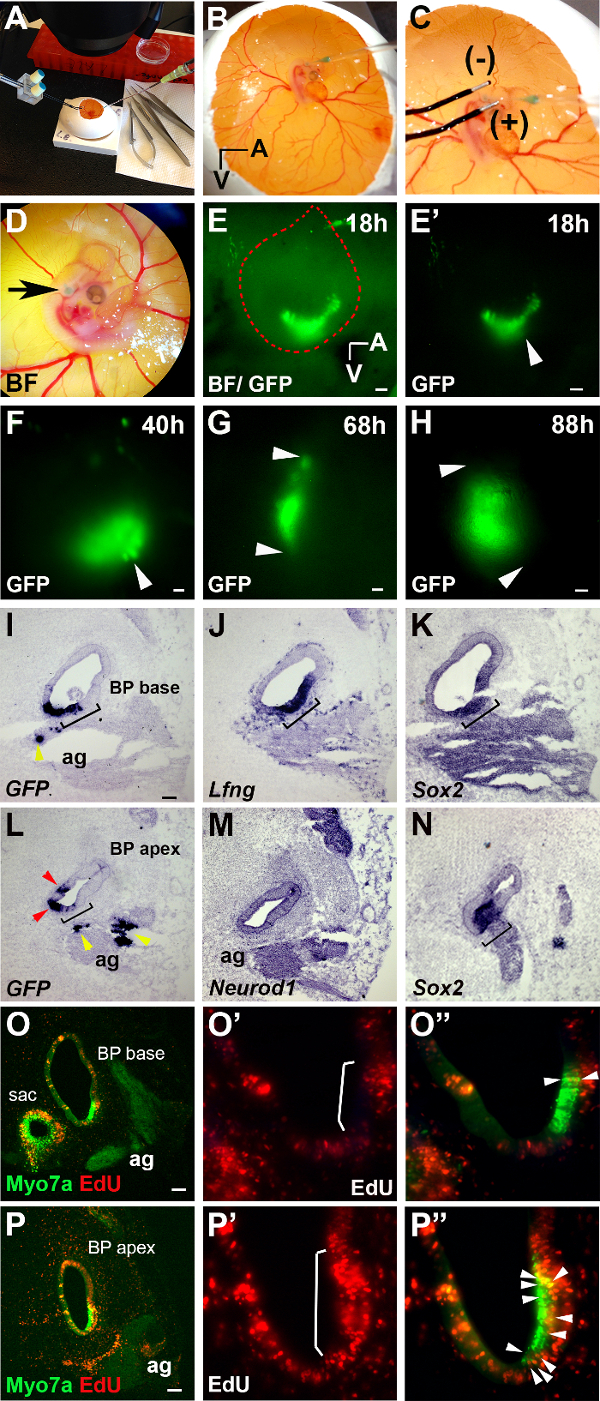
Figure 1. In Ovo Electroporation & In Ovo Cell Proliferation Assay. (A-D) Brightfield images of electroporation method. In ovo micro-injection into the right otic vesicle (B) and placement of electrodes after micro-injection targeting the anterior-ventral area in the otic vesicle (C). Embryo at stage HH24-25 after micro-injection and electroporation (D; green otic vesicle, black arrow). (E-H) Green fluorescent protein (GFP) expression after electroporation. (E) Brightfield image and GFP expression in the otic vesicle (outlined in red) at 18 hr. (E') GFP expression at 18 hr (otic vesicle, white arrow). (F-H) White arrows point to GFP expression at 40 hr (F, otic vesicle), 68 hr (G, BP), and at 88 hr (H, BP). (I-N) Adjacent cochlear tissue-sections of the BP probed for GFP, Lfng,Sox2, and Neurod1 transcripts by RNA in situ hybridization at 68 hr. Lfng (J, black bracket) and Sox2 (K and N, black bracket) mark the sensory epithelium and Neurod1 (M) marks the auditory ganglion (ag). GFP is expressed within sensory (I and L, black bracket) and non-sensory tissue (I and L, red arrows), and the auditory ganglion (I and L, yellow arrows). (O-P'') Fluorescent images of MyosinVIIa (Myo7a)/EdU labeling. (O'-O'' and P'-P'') Higher magnifications of O and P. (O' and P') More EdU labeling is present within the sensory epithelium at the apex (P', white bracket) than at the base (O', white bracket). O'' and P'') Merged fluorescent images, white arrows point to Myo7a(+)/EdU(+) hair cells at the apex (P'') and the base (O''). More EdU(+) cells are present within the auditory ganglion at the apex (P, ag) than at the base (O, ag). (sac; saccule in O). (A, Anterior and V, Ventral in panels B and E). Scale bars 100 µm. Please click here to view a larger version of this figure.
