The feeding system described above can be used for long term manipulation of rhythmic food intake in mice. This system essentially exposes a new food compartment to the mouse every 3 h enabling the researcher to specifically manipulate food in every compartment. One application was to analyze the profile of food intake over the 24 h period. The data indicate that WT mice fed normal chow ad libitum eat about 75% of their food during the night (Figure 2A). Moreover, most of the food eaten during the day occurs within the 3 h before light off.
Mice fed HFD ad libitum ate more food on the first 2 days of exposure, likely because of the novelty of HFD (Figure 2A). After 2 days, the HFD intake remained rhythmic, yet with a decreased amplitude compared to when fed normal chow ad libitum. While both male and female WT mice were fed HFD, it was found that female mice hoarded a large amount of food on the lid of the feeding apparatus and in the cage, whereas males did not show any noticeable hoarding. As mentioned above, hoarding food can result in miscalculating food consumption and lead to data misinterpretation. In addition, female mice bit into the plastic rims of the food cups more frequently, especially in the night compartments. Male mice showed significant weight gain after 1 week of ad libitum normal chow and after 1 week of HFD (Figure 2E). A similar trend was observed with female mice but did not reach significant p-values, likely in part because of the lower number of females used compared to males.
Mice transitioned to a NR diet eat their daily total calories only at night, without a significant decrease in calorie intake for the first 3-5 weeks (Figure 3A). Longer exposure to NR schedule decreases the daily average calorie intake by 10%-15% compared to mice fed ad libitum, as described elsewhere34. Mice transitioned to an AR diet consumed their daily total calories in equal quantities across the day, leading to a dramatic dampening of the daily rhythm of food intake (Figure 3B). As for the NR feeding schedule, the daily average of calorie intake is not affected by the AR feeding schedule for the first 3-5 weeks of exposure but decreases with longer exposure. Mice showed weight gain after the NR (Figure 3C) and AR schedules (Figure 3C).

Figure 1: Design and construction of the feeding apparatus system. (A) The dimensions of the PVC base for the feeding system, and description of where holes need to be made for fixing the timer. (B) A 24 h timer before and after repurposing the wire and drilling with screws for placement of the food cup. (C) The assembled gray base and timer along with a 4-inch PVC pipe. (D) The eight-compartment food cup after trimming outer edges. (E) The final setup of cages with the food cup covered with a 4-inch cap such that only one compartment is accessible at a time. (F) The transportation of multiple food cups during an experiment. Please click here to view a larger version of this figure.
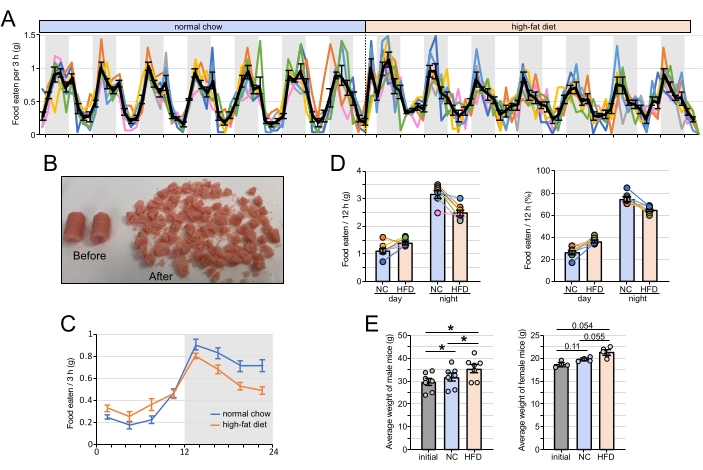
Figure 2: Feeding profiles under different dietary regimes. (A) The feeding profile of male WT mice fed ad libitum with normal chow (NC) for 7 days and high-fat diet (HFD) for an additional 7 days. Colored lines represent individual mouse profiles (n = 7) and the black line indicates the average ± SEM of seven mice. (B) The high-fat diet before and after slicing. (C) The daily average of food intake every 3 h ± SEM (n = 7). The average was calculated over the last 5 days of either the NC or the HFD feeding schedule. (D) The average (left) and the percentage (right) of food intake during the day and night for mice fed with NC or HFD. The values represent the average of seven mice ± SEM and were calculated using the food intake data over the last 5 days of either NC or HFD feeding schedules. * p < 0.05 between the two groups (paired t-test). (E) The average body weights of mice used in the experiment after 1 week of NC and 1 week of HFD. The data for males (left) and females (right) is shown with * p < 0.05 between the two groups (paired t-test). Please click here to view a larger version of this figure.
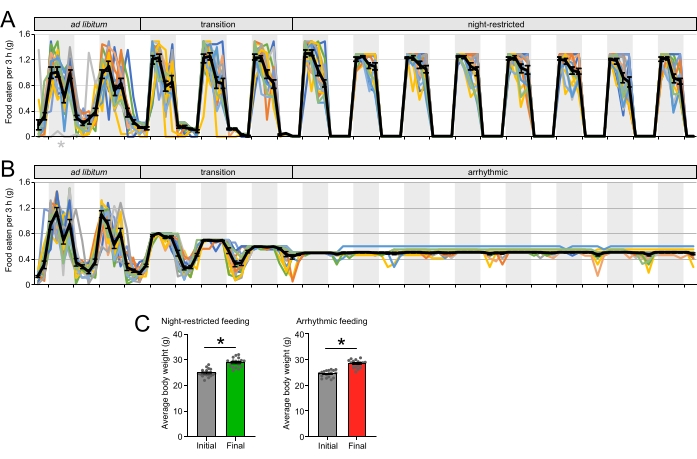
Figure 3: Manipulation of the daily rhythm of food intake. (A) The feeding profile of male WT mice fed with normal chow ad libitum for 2 days, transitioned to a night-restricted (NR) feeding regime for 3 days, and maintained under NR feeding for 8 nights. The colored lines represent individual mouse profiles (n = 18) and the black line indicates the average ± SEM of 18 mice. The gray asterisk indicates malfunctioning of the timer for that mouse on that single day where the timer stopped turning. (B) Feeding profile of male WT mice fed with normal chow ad libitum for 2 days, transitioned to an arrhythmic (AR) feeding regime for 1 day, and maintained under AR feeding for 8 nights. The colored lines represent individual mouse profiles (n = 18) and the black line indicates the average ± SEM of the 18 mice. (C) The average body weights of mice used in the experiment after 2 weeks of exposure to NR and AR diets. Data is shown with * p < 0.05 between the two groups (paired t-test). Please click here to view a larger version of this figure.
Table 1: Advantages and disadvantages of the existing feeding systems. A table highlighting different feeding systems used for manipulation of food intake, with a brief description of the pros and cons of each system. Please click here to download this Table.
Table 2: Cost of materials required to construct the feeding system. A table listing the cost of items required for the construction of the feeding system described in this paper along with an estimate of the cost of construction per cage. Please click here to download this Table.
