Of 34 (n = 18 males, n = 16 females) animals from six litters exposed to the insult, eight animals (24%; n = 4 males, n = 4 females) in the injured group died during the second hypoxia period (n = 5), during temperature management (n = 2), or overnight after the insult (n = 1). In the injured group, nine of 26 survivors (35%) had visible gross injury. Five animals (n = 5 males) had moderate injury, and four animals (n = 2 males, n = 2 females) had severe injury, defined as gross pathology scores of 2–5 and 6–9, respectively (Figure 2A). Animals exposed to the insult therefore have a 50% risk of death or significant gross injury. With increasing injury, narrowing of gyri in the temporal and/or occipital lobes is seen, with associated sulcal shortening, widening of the longitudinal fissure, and large areas of areas of cystic tissue loss in the most severely-injured animals (Figure 2B). In surviving injured animals (n = 26; n = 14 males, n = 12 females), significantly greater exposure of the cerebellum is seen (Figure 3A), as well as shortening of the longitudinal fissure (Figure 3D). There is also significant narrowing of the coronal and anterior ectosylvian gyri (Figure 3B,E), as well as shortening of the lateral and suprasylvian sulci (Figure 3C,F). Median (IQR) brain weight was 8.1 g (7.9–9.7g, n = 6) in control animals, and 7.0 g (6.5–7.7 g) in injured animals (n = 26, p = 0.005). In control animals, median (IQR) brain length was 28.9mm (27.8–29.6 mm, n = 6) compared to 27.5 mm (25.5–38.0 mm, n = 26) in injured animals (p = 0.007). Similar patterns are seen across the brain, with median width and height 5–7% smaller in injured animals. Anatomical structures on both the left and right side are affected in a similar manner, with no difference between hemispheres. See Figure 1B for depictions of the anatomical locations. Over the reflex testing period (P21–P39), injured animals display slower time to rotate in the negative geotaxis task (Figure 4A), slower time to rotate away from the edge in the cliff aversion task (Figure 4B), and slower time to right (Figure 4C). In the catwalk, injured animals have a similar average speed to controls (Figure 5A), but display a significantly greater degree of speed variation during each run (Figure 5B). The weight-adjusted distance between fore paws and hind paws (print position) is significantly greater in injured animals (Figure 5C), with less pressure exerted per unit paw area through the fore paws (Figure 5D). In the open field, injured animals cover less total distance (Figure 6A), and stop more frequently (Figure 6B). They spend significantly more time in the center of the field, and less time in the corners (Figure 6C,D). Representative heat maps of control and injured animals are shown in Figure 7A,B.
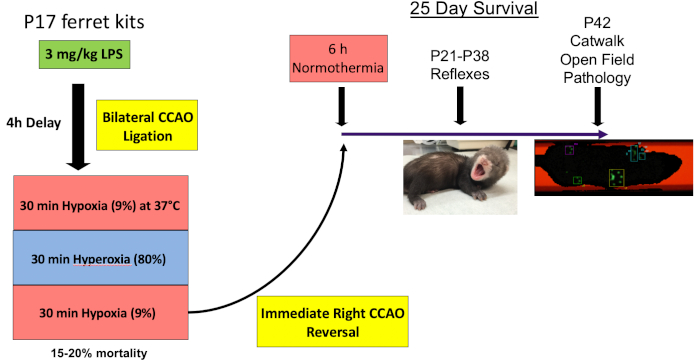
Figure 1: Timeline. On P17, animals are administered 3 mg/kg LPS before undergoing bilateral carotid artery ligation and 30 min each (not including time for the chamber to equilibrate) of hypoxia (9% oxygen), hyperoxia (80% oxygen) and hypoxia (9%). The right carotid artery ligation is then reversed. Animals are exposed to 6 h of normothermia to ensure they do not become spontaneously hypothermic in the nest in the period after injury. Reflex testing is then performed daily from P21–P28, and three times per week from P28–P42. On P42, animals are tested in the catwalk and open field before sacrifice. Please click here to view a larger version of this figure.
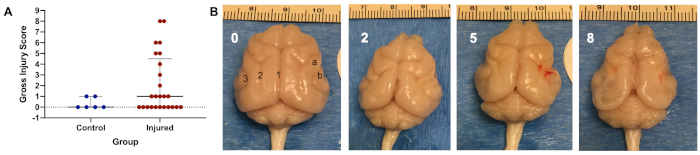
Figure 2: Representative injury distribution and depiction. (A) Gross injury scoring from 26 survivors (n = 14 males, n = 12 females) in the injured group, compared to six litter mate controls. Five animals (n = 5 males) had moderate injury, and four animals (n = 2 males, n = 2 females) had severe injury, defined as gross pathology scores of 2–5 and 6–9, respectively. Graph shows median with interquartile range. (B) Control brain (left panel, score 0), with brains depicting increasing gross injury scores of 2, 5, and 8 out of a total possible score of 9, from left to right. The control brain shows anatomical structures particularly susceptible to injury; 1 = longitudinal fissure, 2 = lateral sulcus, 3 = suprasylvian sulcus, a = coronal gyrus, b = anterior ectosylvian gyrus. Please click here to view a larger version of this figure.
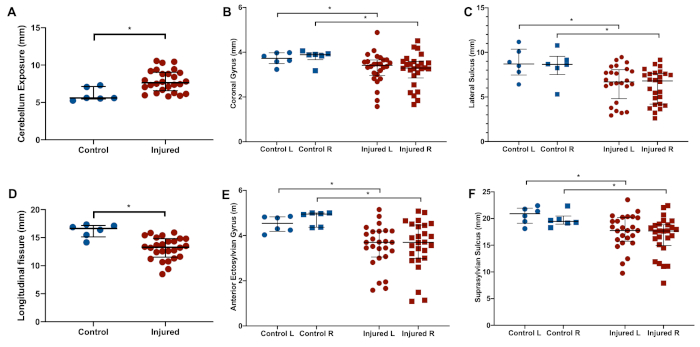
Figure 3: Representative brain measurements. Compared to controls (n = 6), injured animals (n = 26) display significantly increased exposure of the cerebellum (A), shortening of the longitudinal fissure (D), narrowing of the coronal (B) and anterior ectosylvian (E) gyri, and shortening of the lateral (C) and suprasylvian (F) sulci. Graphs show median with interquartile range. *denotes p < 0.05 (Wilcoxon-Mann-Whitney U-test). Please click here to view a larger version of this figure.
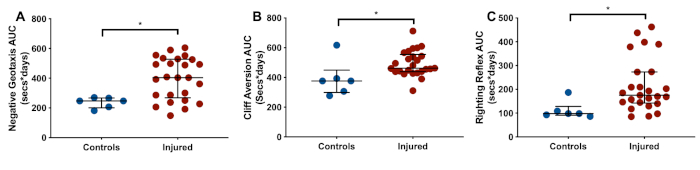
Figure 4: Representative reflex development. Compared to controls (n = 6), injured animals (n = 26) display slower development (area under the curve, AUC) of negative geotaxis (A), cliff aversion (B), and righting reflex (C). Graphs show median with interquartile range. *denotes p < 0.05 (Wilcoxon-Mann-Whitney U-test). Please click here to view a larger version of this figure.
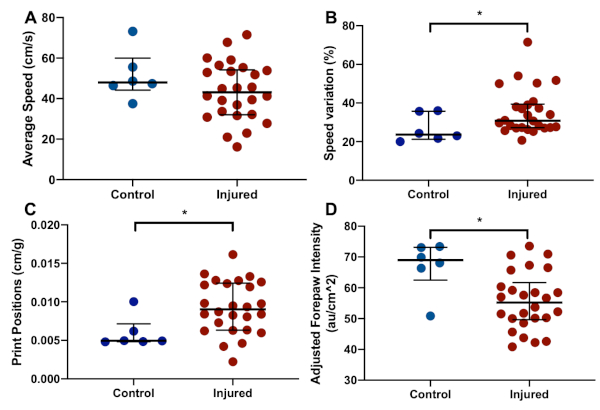
Figure 5: Representative catwalk results. Compared to controls (n = 6), injured animals (n = 26) walk at a similar average pace (A), but with a greater variability in speed during walking (B). Injured animals also display a longer average print position (C), with less pressure applied per unit area (D). Graphs show median with interquartile range. * denotes p < 0.05 (Wilcoxon-Mann-Whitney U-test). Please click here to view a larger version of this figure.
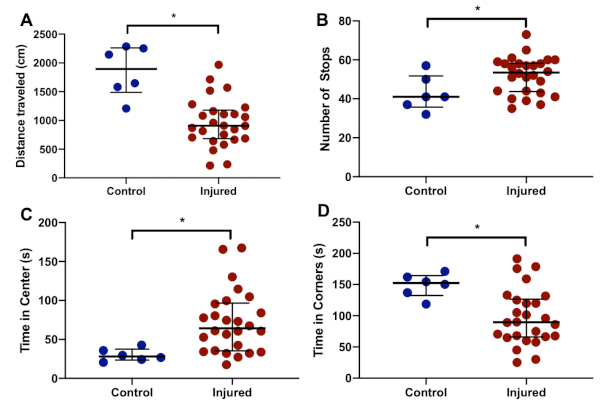
Figure 6: Representative open field behavior. Compared to controls (n = 6), injured animals (n = 26) cover a smaller total distance (A), as well as stopping more frequently (B). Injured animals also spend more time at the center (C) than in the corners (D). Graphs show median with interquartile range. *denotes p < 0.05 (Wilcoxon-Mann-Whitney U-test). Please click here to view a larger version of this figure.
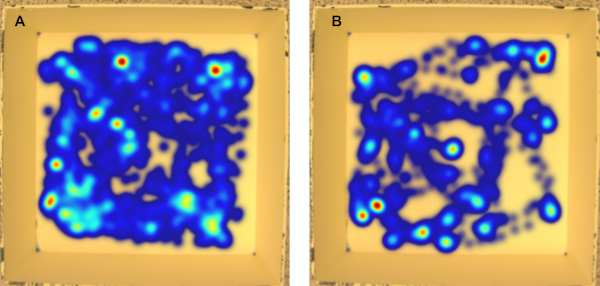
Figure 7: Representative open field heat maps. (A) control female, (B) injured female. Injured animals cover a significantly smaller distance within the open field. Please click here to view a larger version of this figure.
