NP-exposed cells were processed with the cell staining reagent kit, followed by cell sorting using flow cytometry. ZnO NP-treated cells (bottom, right panel) exhibit a higher percentage of early (R3)/ late apoptotic cells (R6) than control cells (R5, bottom, left panel). Necrotic cell death is denoted by R4 (top, right panel) (Figure 2). The results of the FITC/Annexin V Assay on ZnO NP-treated MRC-5 fibroblasts are shown in Figure 2.
For the Drosophila experiments, sonicated ZnO NPs at various concentrations were added to fly food in 10 mL tubes and then mixed well using a pipette controller (Figure 3). Late third instar larvae collected from vials were dissected under the stereomicroscope. The larvae were first washed to remove remnants of remaining food (Figure 4). The outer cuticle layer was torn apart to expose internal organs. The gut was identified by the characteristic long and semitransparent appearance (whereas other organs appear opaque and light yellowish under the microscope) (Figure 5). The gut was carefully removed, without breaking, and transferred into a new microcentrifuge tube containing fixative on ice (Figure 6).
For the quantitation of fluorescence intensity such as the intensity of the DHE probe in the gut, the images were exported in JPEG or TIFF formats and opened with the ImageJ software. The part of the gut for analysis was selected and identified, for example, the midgut or hindgut region, and the fluorescence intensity of the region of interest (ROI) was measured. To compare the relative intensities of the different experimental groups, we employed the same quantitative confocal microscopy method described in the previous section. For comparison of fluorescence intensities, the parameter was set using the negative untreated control. Calculations of the signal intensity on the basis of calibration intensities of untreated control allowed a direct comparison between different experimental groups. Figure 7 shows the average intensity of the DHE signal in the 3rd instar larval gut exposed to ZnO NPs at different concentrations. The gut of larvae treated with 0.5 mg/mL of ZnO NP treatment showed the highest fluorescence intensity.
The differences in relative intensities between all the experimental groups were further tabulated, and statistical analysis was performed, providing both qualitative and quantitative results (Figure 8).
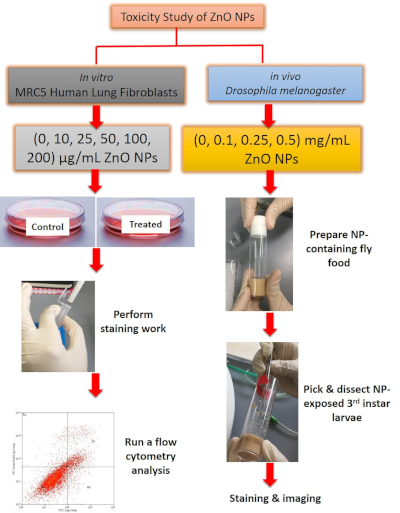
Figure 1: Overview of the method used for toxicity study of ZnO NPs. For in vitro work, ZnO NP-treated cells were stained prior to flow cytometry analysis. For in vivo work, gut was dissected from 3rd instar larvae, followed by staining with DHE dye and image acquisition. Please click here to view a larger version of this figure.
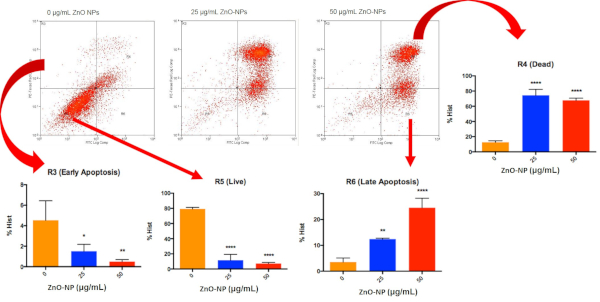
Figure 2: Dot plot of cells separated into different populations based on their FITC and PE staining. The pictograms show the results of FITC/Annexin V assay with 24 h treatment of ZnO-NPs on MRC-5. Statistical analysis of the cells at different stages can then be performed. Please click here to view a larger version of this figure.

Figure 3: Preparation of ZnO NP-containing fly food medium. (A) Ingredients for fly food are added to water, allowed to swell, and boiled for 5 min. (B) After cooling down to 50 °C with stirring, Nipagin was added and mixed thoroughly. (C) Prepare a master mix for the nanoparticles (total volume not exceeding 10% of the final food volume). (D) Medium is then aliquoted, mixed with ZnO NPs at various concentrations and allowed to cool down completely before storage. Please click here to view a larger version of this figure.
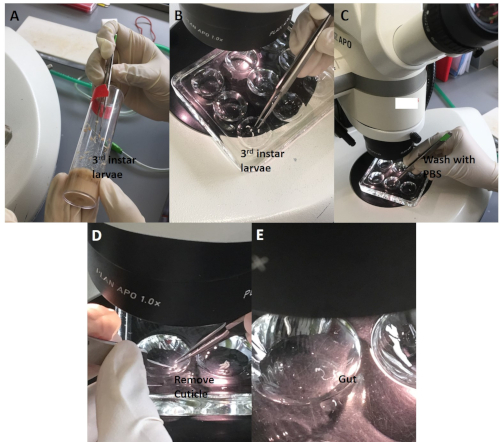
Figure 4: The whole gut dissection procedure. (A) Transfer 3rd instar larvae to a dissection disc. (B) Use forceps to gently hold a larva, and (C) wash away the remnant of food using saline. (D) Gently tear the cuticle apart without touching the gut and other internal organs. (E) Place the gut into the saline for subsequent procedure. Please click here to view a larger version of this figure.
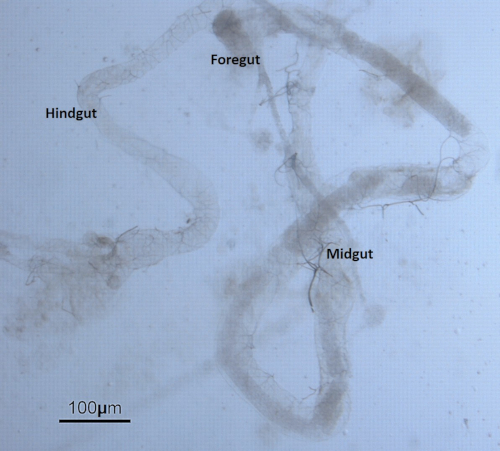
Figure 5: The anatomy of the digestive tract/gut. The gut extracted from the Drosophila larva is divided into three discrete domains of different developmental origin namely the foregut, midgut, and hindgut. Please click here to view a larger version of this figure.

Figure 6: Staining of the gut tissue with DHE. (A) Prepare a master mix containing growth medium and DHE (a final concentration of 30 µM). (B) Add the master mix into the well of a dissection disc. (C) Transfer the dissected gut tissue into the well containing the DHE master mix. (D) Incubate at RT for 5 min and protect the tissue from light; wash 3x in PBS/saline for 5min. (E) Fix in 4% PF for 10 min; wash the tissue three times with PBS (optional). (F) Gently transfer the gut onto a glass slide, lay flat without having any tissue folded and mount with mounting medium before covering with cover glass. Please click here to view a larger version of this figure.
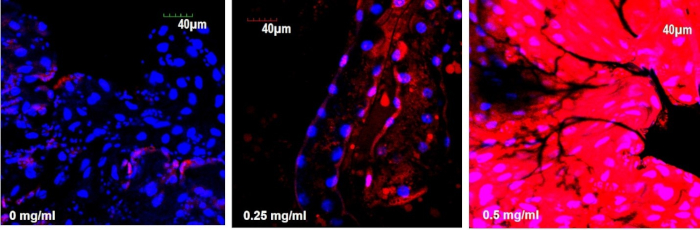
Figure 7: ZnO NPs induce ROS in the gut of Drosophila larvae. (a) DHE stain in control gut shows the basal level of ROS. (b and c) Treated gut cells show a gradual increase in DHE intensity in a dose-dependent manner. Please click here to view a larger version of this figure.
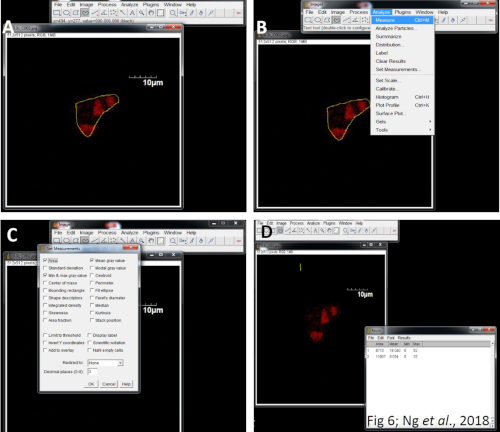
Figure 8: Quantitation of fluorescent images using ImageJ. (A) Import the captured images. (B) Click on the Analyze menu and select Set measurements. (C) Select the area integrated intensity and mean grey value. Select a region without fluorescence to set the background. (D) Export the data into Excel and calculate the CTCF for subsequent statistical analysis. Please click here to view a larger version of this figure.
