The manuscript describes an optimized protocol to easily monitor the behavior of monocytes and neutrophils in the mesenteric veins of anaesthetized mice in real time. The use of a 37 °C-thermostated chamber is mandatory to maintain the temperature of the mouse and also due to the temperature dependent movement of leukocytes. Preparation of the mouse is displayed in Figure 1. Figure 2 shows all the area seen under the microscope. Transmitted light allows the identification of mesenteric veins (red arrow) and arteries (blue arrow).
Treatment of acquired images with ImageJ software generated movie files, such as Movie 1 and Movie 2. Figure 3 displays the different steps of image processing for the first time point of Movie 1. Movie 1 shows the patrolling of Ly6Clow monocytes under steady state conditions, as previously described 6,8,14 whereas all neutrophils are circulating in the bloodstream. No neutrophil is patrolling the endothelial wall. Movie 2 shows the same field of interest 90 min after the addition of the TLR2/TLR1 agonist Pam3CSK4 to locally initiate inflammation. In this case, neutrophils and monocytes are massively recruited to the endothelial wall, which they scan meticulously. It is important to note that addition of a TLR agonist induces changes in endothelium width and consequently alters the focus of the z-axis. Therefore, focus should be carefully controlled after the addition of the agonist. Compilation of data in Imaris software (Bitplane) allows the tracking of individual leukocytes using the Spot Tracking Wizard. Figure 4 shows the exact track paths of monocytes (green) and neutrophils (red) patrolling along the endothelium. The data are obtained from the analysis of Movie 2, i.e., 90 min after the initiation of inflammation. Tracking of leukocyte paths can be exported as tiff files or xls files to get various statistics such as track length, displacement, duration or speed.
This technique offers a nice way to monitor the behavior of monocytes and neutrophils at the same time before and after inflammation. It is possible to track the recruitment of these cells on the endothelial wall overtime. Loss of focus on the z-axis or even x/y displacement can occur as a consequence of intestinal movements that ruin the experiment. Movie 3 illustrates such failed preparation and acquisition.
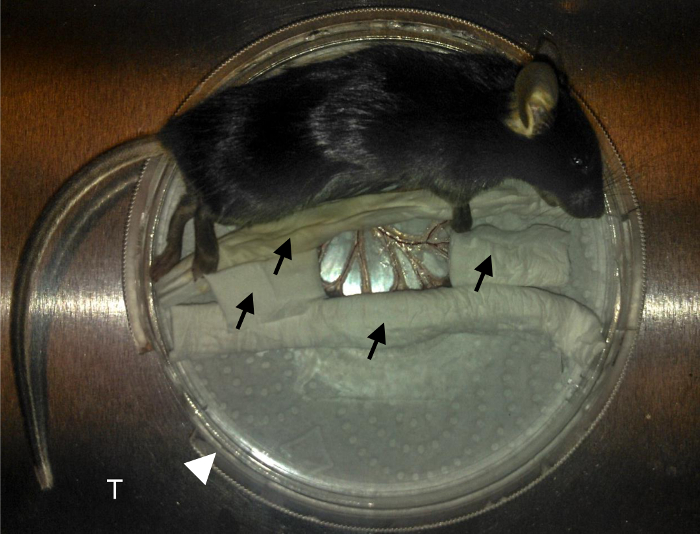
Figure 1. Mouse preparation.
Black arrows indicate PBS-wetted tissue used to immobilize the intestine. White arrowhead indicates the 10cm tissue culture dish. T indicates the aluminum custom-made tray stage that fits into the microscope. Mesenteric blood vessels are nicely exposed at the center on the coverslip. Please click here to view a larger version of this figure.
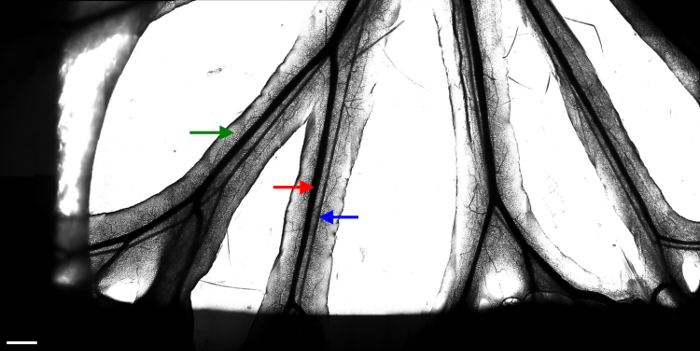
Figure 2. Branches of the mesenteric veins and arteries.
Images (20 x 10) taken with a 10X objective were stitched to constitute one large image of the available area. Several fields of interest are then chosen to monitor leukocyte movement with a 20X objective. The red arrow indicates a mesenteric vein. The blue arrow indicates a mesenteric artery. The green arrow indicates the fat tissue surrounding the vessels. The dark area on the sides of the image results from the presence of wetted tissue used to immobilize the intestine. White scale bar represents 1 mm. Please click here to view a larger version of this figure.
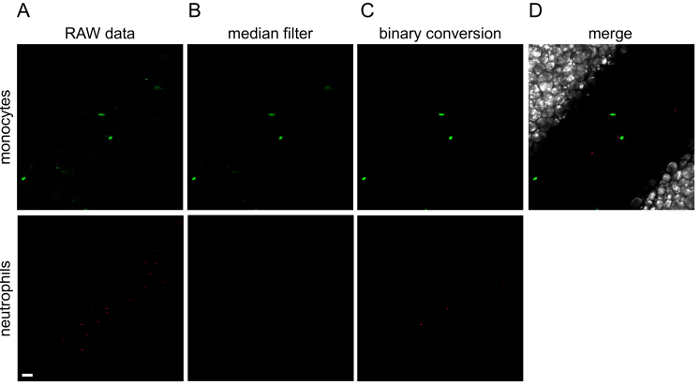
Figure 3. Image processing with ImageJ.
The different steps of image processing are illustrated for the first image of Movie 1. Green (monocytes) and red (neutrophils) channels are processed separately. To the original raw data (A), a median filter is applied (B) and then converted to a binary image (C). (D) Channels are merged into a final image. Green or red lines observed in raw data are cells that are moving so fast that the microscope is not able to wholly acquire them. White scale bar represents 40 µm. Please click here to view a larger version of this figure.
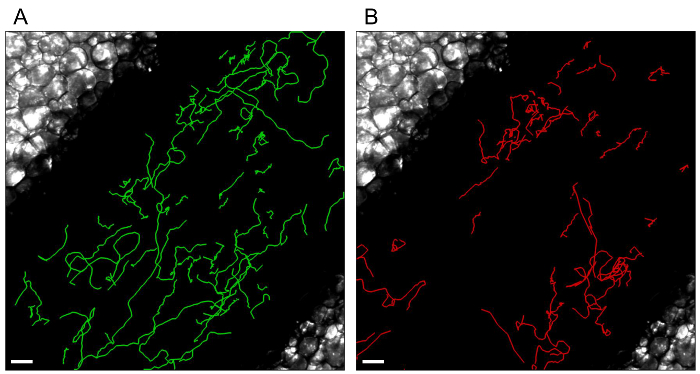
Figure 4. Individual leukocyte tracks.
Tracks of monocytes (A) and neutrophils (B) patrolling the endothelium are processed with Imaris Software (Bitplane). White scale bar represents 40 µm. Please click here to view a larger version of this figure.
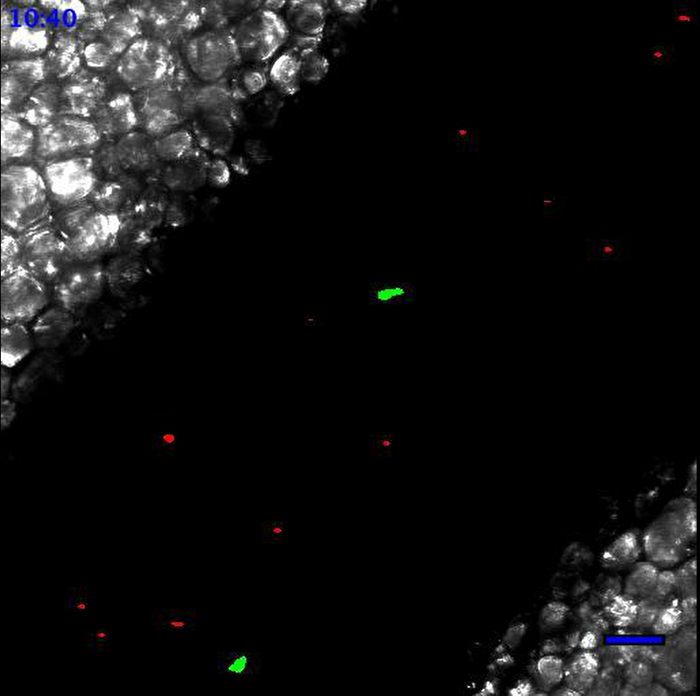
Movie 1. Leukocytes scanning the endothelial wall at the steady state. At the steady state, Ly6Clow monocytes patrol the endothelium. Neutrophils are circulating according to the blood flow. Neutrophils are in red and monocytes are in green. Scale bar represents 50 µm. Generated with the 20X objective. Please click here to view this video.
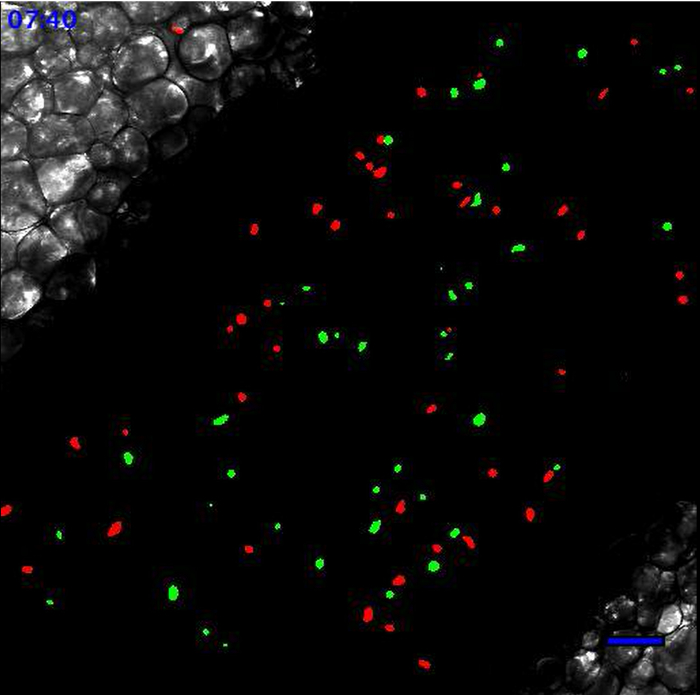
2: Leukocytes scanning the endothelial wall after TLR2-mediated inflammation. Same field of interest as in Movie 1. Movie starts 90 min after the addition of 20 µl of PBS containing 100 µg of Pam3CSK4 (TLR2/TLR1 agonist) directly onto the vessel. Recruited monocytes and neutrophils are meticulously scanning the endothelial wall. Neutrophils are in red and monocytes are in green. Scale bar represents 50 µm. Please click here to view this video.
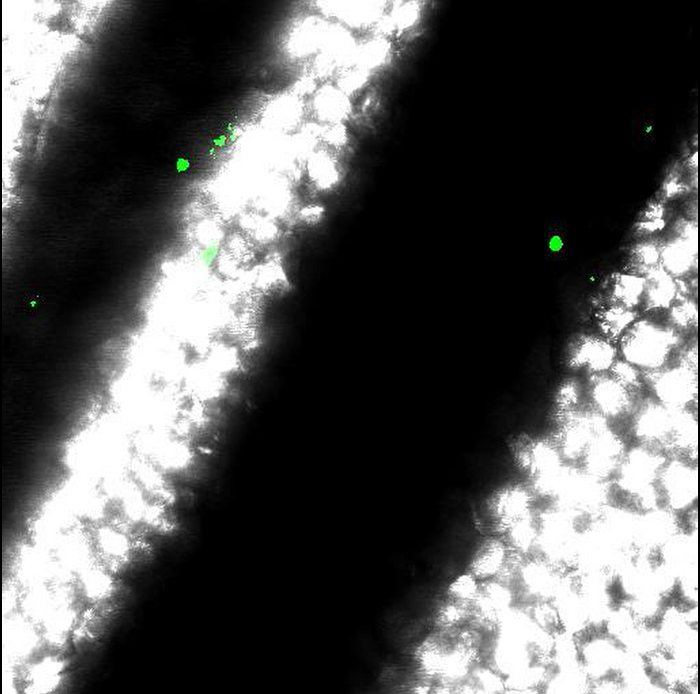
Movie 3: Results obtained from incorrect mouse preparation. Movements of vessels obtained when anesthesia or immobilization of intestine are incorrect. Scale bar represents 50 µm. Please click here to view this video.
