It has become clear over the past several years that the complexity of the phloem sap may be the answer to the question of how plants send varying developmental and stress signals for optimal response. Using EDTA-facilitated phloem exudation may provide the opportunity to analyze phloem exudates from plants that are not suitable for other approaches but may be of economic or physiological interest.
This method allows for the harvest of enough phloem exudates to identify phloem-localized proteins and detect changes in their level in response to development or stress (Figure 3). As the figure shows, proteins are in very low abundance, yet, their level is high enough for subsequent proteomics experiments using LC-MS/MS or for Western blot analysis. Findings in cucurbits suggest that the cutting of the stem leads to a disruption of the water potential balance and a subsequent influx of water and possible contaminants from the apoplast41. Yet collection for times ranging from one to eight hours does not affect the phloem protein profile/composition in Arabidopsis (only the absolute amount varies Guelette et al.), the amount of protein collected and the pattern of proteins found varies between species (Arabidopsis: A.t.; Perilla, lane I and NI) and treatments (Perilla grown at different day lengths, lane I and NI). As a result proteinaceous signals can be visualized, identified, and followed.
mRNA and micro RNAs can also be identified in phloem exudates by this method. However, treatment with RNAse inhibitor is necessary to prevent degradation of the mRNA during the extended exudation time. Since the sieve elements do not contain functional chloroplasts, Rubisco small or large subunit (RbcS or RbcL, respectively) mRNAs can be used as negative controls, while known phloem-localized mRNA like Ubiquitin-conjugating enzyme may be used as a positive control (Figure 4).
Similarly, sugars or small metabolites can be analyzed using either GC-MS or LC-MS. Here it should be mentioned that the phloem exudates contain a large number of functional enzymes, including almost the entire glycolytic pathway12. Hence, using several time points may reveal metabolic processes during the exudate collection. An example is shown in Figure 5: In all exudates, sucrose is the most abundant metabolite; this is most obvious after a collection for 1 hour. However, after five hours the sucrose to fructose ratio is slightly reduced. If the same exudate is collected for one hour and left on the bench for the next four hours, the sucrose to fructose ratio is reduced to a much larger extent, suggesting that active enzymes in the phloem exudates lead to a degradation of sucrose at room temperature (for more peak interpretations see 14). This is consistent with findings that phloem loading and transport of molecules from companion cells into the sieve elements may continue during exudation until the system loses its vitality34.
Lipids in the phloem exudates and, by extension, long-distance lipid signaling are a fairly recent field of interest in plant science. Due to their hydrophobic nature lipids are only present in low concentrations and may be bound to other molecules for solubilization. Yet, the EDTA-facilitated exudation allows for the collection of sufficient material to visualize (TLC; Figure 6) and identify (LC-MS; Figure 7) phloem lipids from several plant species. As the figure shows it is possible to identify and separate several lipid species. LC-MS allows to identify different lipid species and to monitor changes within the lipid profiles of different genotypes or treatments. This allows for the study of the role of lipids during plant development and stress response.
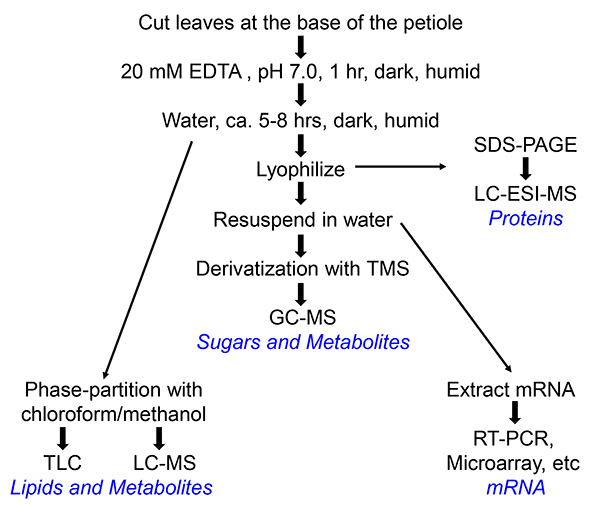
Figure 1. Flow chart of the collection of phloem exudates of Arabidopsis or Perilla. Different paths lead to different end products, which are indicated in blue. For preparation of RNA, RNAse inhibitor (100 U/ml, Roche) is added to the water into which the phloem exudate is collected.
Click here to view larger image.

Figure 2. Setup of materials for phloem exudate collection. The setup displays all materials needed for protocol steps 3.1-3.4.
Click here to view larger image.
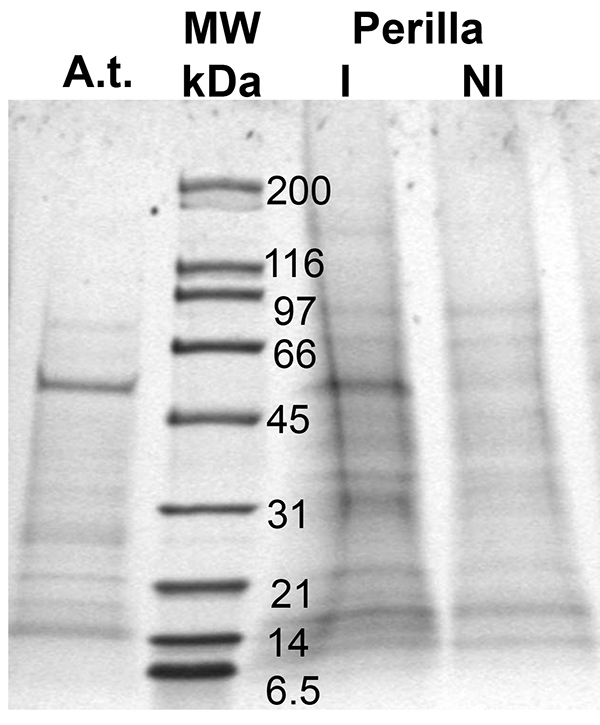
Figure 3. Proteins found in phloem exudates of two different plant species, Arabidopsis (A.t.) and Perilla (I and NI; different day lengths); MW: molecular weight marker (lane 2). Proteins were separated using a 10-20% gradient SDS-PAGE.
Click here to view larger image.
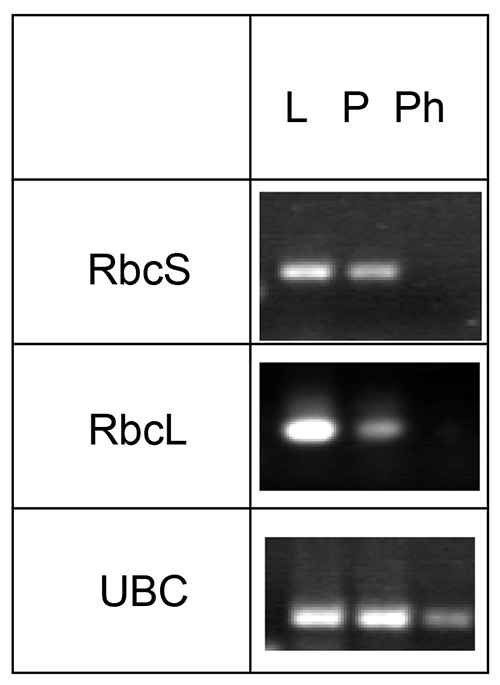
Figure 4. Analysis of the presence of mRNA for Rubisco small and large subunit (RbcS and RbcL, respectively) and ubiquitin-conjugating enzyme (UBC). mRNA was collected from Arabidopsis leaves (L), petioles (P), and phloem exudates (Ph), reverse transcribed and specific transcripts visualized using PCR. The figure was modified from Guelette et al.14.
Click here to view larger image.
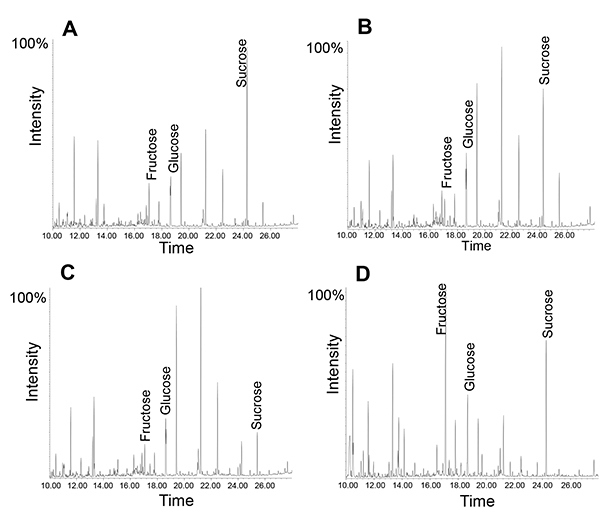
Figure 5. GC-MS profile of phloem exudates collected for different amount of times. Note how the relative amount of sucrose changes from 1 hour collection (A) to 5 hours of collection time (B). Exudate profile after collection for 1 hour followed by "incubation" at room temperature (RT) for four hours (C). Leaf metabolite profile (D).
Click here to view larger image.
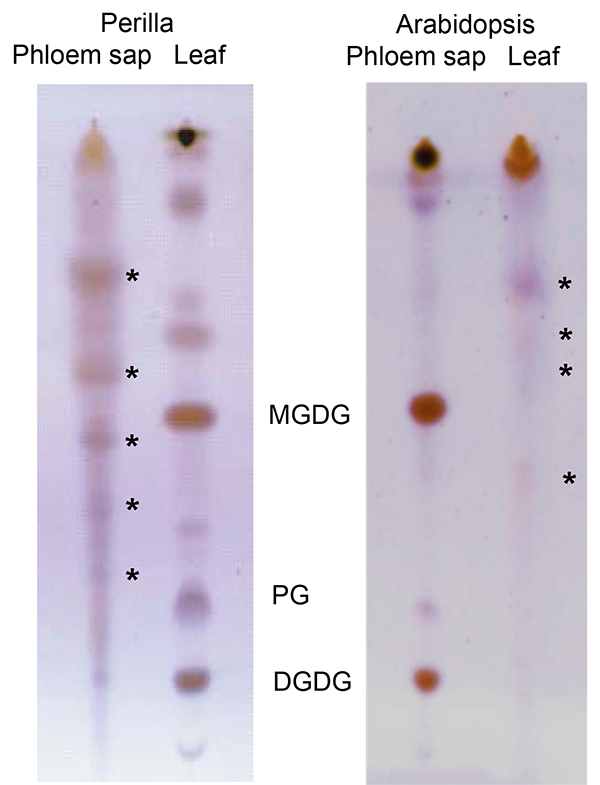
Figure 6. Thin-layer chromatogram comparing lipids from Perilla (left) and Arabidopsis (right) leaves and phloem exudates. Asterisks indicate lipids specific for phloem exudates. DGDG: digalactosyldiacylglycerol; PG: phosphatidylglycerol; MGDG: monogalactosyldiacyglycerol; The part of the figure displaying Arabidopsis lipids has been modified from Guelette et al.14.
Click here to view larger image.
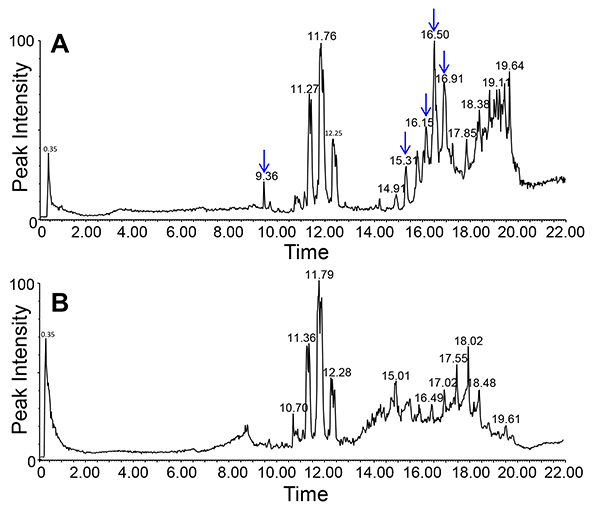
Figure 7. Lipid profile of phloem exudates of Arabidopsis (chloroform phase) using LC-MS/ negative ion mode. Graphs in the top (mutant, A) and bottom (wild-type, B) show the differences in the lipid profiles between two different genotypes (indicated by arrows).
Click here to view larger image.
