The transuroepithelial neutrophil migration assay enables the quantitative assessment of human PMN migration across cultured bladder epithelial cell layers in response to various stimuli (Figure 1B). While the protocol is straightforward, there are a number of variables that can influence PMN migration and consequently affect the reproducibility of this assay. Measures should be taken while preparing the permeable supports and the PMN to reduce variability between technical and biological replicates. For example, only permeable supports containing sufficiently confluent 5637 cell layers should be used in an experiment. Confluence of the 5637 cells is assessed using a functional assay that measures impermeability to liquid. If medium added to the upper reservoir equilibrates across the permeable support, then the 5637 cells are not sufficiently confluent to conduct the experiment. If the volume in the upper reservoir is maintained, then the permeable support can be used to assess PMN migration. We have measured transepithelial electrical resistance in this system, which rises modestly upon confluence of the cells; if this method is chosen, care should be taken not to contaminate the otherwise sterile setup. Confluence of the 5637 cells 7 days after seeding can be influenced by multiple factors, including the passage number of the cells and the number of cells seeded on the permeable support. In addition, the amount of time that the 5637 cells are incubated on the permeable support in the inverted position during seeding should not exceed 16 hr (Figure 1A). For optimal reproducibility, the protocol should be followed precisely. Finally, permeable supports containing confluent 5637 cell layers should be used within 1-2 days, and the membranes of the supports should never be touched during either growth of the 5637 cells or during the transuroepithelial neutrophil migration assay.
In addition to the 5637 cells, variability can also be introduced during PMN preparation. Using the protocol detailed above to isolate PMN, 1 ml of human blood typically yields about 106 PMN, although this number varies from individual to individual. Once the typical yield of an individual donor's blood is known, the isolation protocol can be scaled up or down accordingly. PMN from unhealthy or ill individuals should be avoided, and different PMN donors should be used for biological replicates to ensure that results observed are reproducible. PMN should be handled gently and aseptically to avoid activation during isolation. Lastly, the timing of the experimental procedures is crucial, as PMN do not survive for extended periods of time once removed from the body. We utilize PMN within 1 hr of completing the isolation procedure. Given these considerations, at least 3 technical replicates should be included in each biological replicate.
The number of PMN in the lower reservoir after 1 hr is shown in (Figure 2) normalized to 106 input PMN. Alternatively, PMN numbers can be compared to an internal control after normalization to input PMN, which may reduce variation between biological replicates. Adherence to the protocol outlined above with attention to detail enables the enumeration of PMN migration in response to stimuli including bacteria (Figure 2A) and chemoattractant substances (Figure 2B).
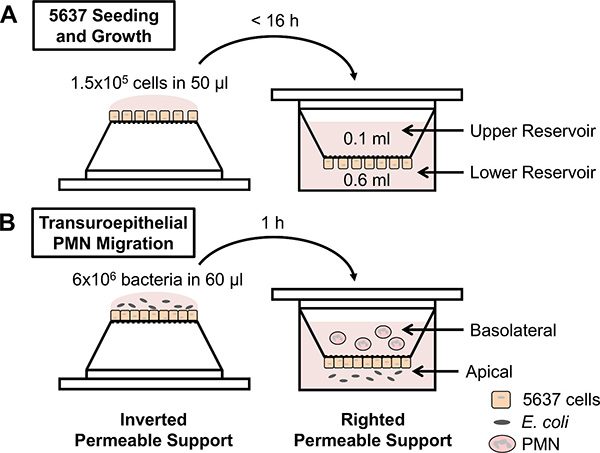
Figure 1. Schematic of experimental design. (A) 5637 bladder epithelial cells are seeded on inverted permeable supports, the supports are righted into a 24-well plate, and the cells are grown to confluence. (B) Permeable supports containing confluent 5637 cells are inverted and infected with E. coli on the apical side of the epithelial layers. Alternatively, chemoattractants can be placed in the lower reservoir. The permeable supports are righted into a low-attachment plate, and freshly isolated human PMN are applied to the upper reservoir (representing the basolateral side of the epithelial layers). PMN migrate across the epithelium and are enumerated from the lower reservoir using a hemocytometer.
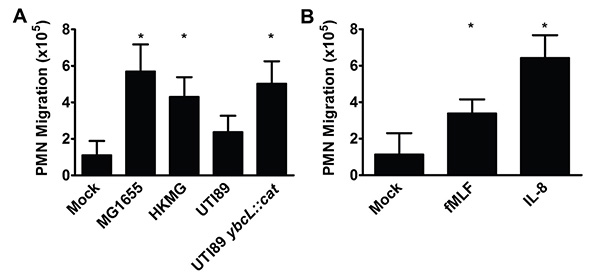
Figure 2. PMN migrate across bladder epithelia in response to various stimuli. (A) Infection with nonpathogenic E. coli strain MG1655, heat-killed MG1655 (HKMG) or UPEC mutant UTI89 ybcL::cat elicits significantly more PMN migration than mock infection or infection with wild-type UPEC strain UTI89, a cystitis isolate (*, p <0.001). (B) The addition of fMLF (100 nM) or IL-8 (100 ng/ml) to the lower reservoir results in significantly more PMN migration than mock treatment (*, p <0.001). Data represent the mean and standard deviation from at least 3 biological replicates. Statistically significant differences were determined using an unpaired Student's t test.
