Purification of FtsZ from different bacterial sources has been described in the literature and is summarized in Table 1.
| Source | Method | Modification | Yield obtained [mg/L of culture]> | Referencias |
| B. subtilis | 1) Ammonium sulfate precipitation/ion exchange chromatography | no | 40 | This work, 11,15 |
| 2) Affinity chromatography | His-tag | ND | 17 | |
| E. coli | 1) Ammonium sulfate precipitation/ion exchange chromatography | no | 35 | This work, 11,15 |
| 2) Calcium precipitation, ion exchange chromatography | no | 40 | 18 | |
| Methanococcus jannaschii | 1) Affinity chromatography under denaturing conditions/refolding/ammonium sulfate precipitation/gel filtration | His-tag | 1.3 | 19 |
| 2) Affinity chromatography/ gel filtration | His-tag | ND | 20 | |
| Thermotoga maritima | Ion exchange chromatography/gel filtration | no | 6.7 | 19 |
| Pseudomonas aeruginosa | Affinity chromatography/gel filtration | Strep-tag, His-tag | ND | 21 |
| Mycobacterium tuberculosis | 1) Affinity/ion exchange chromatography | no | ND | 22 |
| 2) Affinity chromatography/ gel filtration | no | 30 | 23 | |
| Aquifex aeolicus | Affinity chromatography/gel filtration | His-tag, C-terminal truncation (331-367) | ND | 24 |
| Caulobacter crescentus | Ion exchange chromatography/ammonium sulfate precipitation/gel filtration | no | ND | 25 |
Table 1. FtsZ purification protocols described. ND: not determined.
Sedimentation of FtsZ polymers
Initially, we used two different velocities to spin down FtsZ polymers. We found that only at a velocity of 350,000 x g single polymers of FtsZEc are spun down (Figure 1) whereas at 190,000 x g only bundles of FtsZBs are present in the pellet fraction (data not shown). Therefore 350,000 x g was used in our further experiments. The percentage of polymerized FtsZEc and FtsZBs is similar at 50 mM KCl even though the light scattering experiments revealed a much higher scattering signal for FtsZBs. This is due to bundles formed by FtsZBs which scatter more light than single polymers of FtsZEc. It was not possible to obtain high amount of FtsZ polymers in the pellet fraction in the experiment with 300 mM KCl for both FtsZEc and FtsZBs (Figure 1). We attribute this to a combination of quick disassembly of the FtsZ structures and decreased bundling of the filaments.
Sedimentation of FtsZ-SepF tubules
To analyze the interaction of FtsZ with certain activators sedimentation assays can be performed at lower centrifugation speeds. At this velocity only large structures of FtsZ may be pelleted, e.g. the large tubules formed by SepF rings and FtsZBs filaments5, or the bundles formed by FtsZ and ZapA. We used lower centrifugation (24,600 x g) to demonstrate the feasibility of this approach for the tubules formed by FtsZ and SepF. FtsZ was recovered in the pellet above background levels only when both SepF and GTP were present in the sample (Figure 2), and the presence of SepF does not influence FtsZ GTPase activity7 showing that FtsZ is fully active in the presence of SepF. Specific sedimentation of SepF and FtsZ is roughly 45% of total SepF and 15% of total FtsZ (compared to material sedimenting when GDP is added). This shows that the SepF-FtsZ tubules contain more SepF than FtsZ. This may be because many SepF rings organize the FtsZ-SepF tubules5,7. The exact stoichiometry of SepF-FtsZ in these tubules is not known but our results suggest that there is more SepF present than FtsZ.
FtsZEc and FtsZBs polymerization and bundling properties
To characterize the polymerization efficiency of FtsZBs and FtsZEc in different buffers we analyzed both proteins by 90° angle light scattering. At 50 mM KCl, FtsZBs gives a 20-40-fold higher light scattering signal than FtsZEc depending on buffer pH (Figures 3A and B) confirming results of Buske et al.26 Increasing the KCl concentration in the buffer did not significantly influence the light scattering signal of FtsZEc (Figure 3D) but the signal of FtsZBs decreased ~80-fold at pH 7.5, ~30-fold at pH 6.8 and ~45-fold at pH 6.5 in 300 mM KCl (Figure 3C) compared to buffers with 50 mM KCl (Figure 3A). Disassembly of FtsZ polymers is faster at higher KCl concentration for both proteins (Figures 3C and D). Studies of Pacheco-Gómez et al. show that E. coli FtsZ polymerization and bundling is pH dependent. These authors found that in a buffer with 50 mM KCl the light scattering signal of FtsZ polymerization was higher, and disassembly of FtsZ took longer at pH 6.0 compared to pH 7.0 8. These results are not in agreement with our data from polymerization of FtsZEc at 50 mM KCl (Figure 3B), but it has to be noted that we have used three different buffers (HEPES, MES and PIPES) where Pacheco-Gómez et al. only used MES. Thus, not only the pH, but also buffer composition (ionic strength) affects the kinetics of FtsZ polymers at 50 mM KCl. However, at 300 mM KCl neither buffer composition nor pH influenced FtsZEc assembly in a detectable manner.
A light scattering experiment of FtsZBs in buffers without KCl was not possible due to precipitation of the protein under these conditions. When the concentration of FtsZBs was lowered to 3 µM, precipitation did not occur. However, 3 µM is not the physiological concentration of FtsZ in the cell. In plastic cuvettes, FtsZBs did not precipitate at 12 µM at pH 7.5, but at pH 6.8 and 6.5 FtsZ still precipitated in the absence of KCl.
Morphologies of FtsZ structures from E. coli and B. subtilis
The structures formed by FtsZ were inspected by TEM. FtsZBs assembled into closely compacted polymers that covered the whole grid in all buffers at low salt (Figures 4A-C). FtsZEc formed long filaments, cables and bundles in all buffers at low salt (Figures 4D-F). However, the observable amount of polymers formed by FtsZEc was lower than the amount formed by FtsZBs. In high salt buffers FtsZBs formed longer single-stranded protofilaments which did not associate into bundles (Figures 4G-I). While FtsZBs protofilaments changed structure at higher salt concentration, FtsZEc formed structures indistinguishable from those of low salt buffer (Figures 4J-L). There was no observable pH influence on polymerization of FtsZEc but FtsZBs forms more bundles at pH 6.5 which are visible as closely compacted polymers and sheets (Figure 4B). These results are in accord with our light scattering experiments and previously published TEM work8,11,26.
The GTPase activity of FtsZ at high and low KCl concentrations
The GTP hydrolysis activity of FtsZ was measured under different conditions using a colorimetric assay for free phosphate. As reported previously15 the GTPase activity of FtsZ increased with increasing KCl concentration: depending on the buffer used FtsZBs. showed a 3-7 fold increase, and FtsZEc showed a 1.5-2.5 fold increase in GTPase activity at 300 mM KCl compared to 50 mM KCl. The reduced GTPase activity at 50 mM KCl is due to bundling of FtsZBs filaments. At 50 mM KCl FtsZEc had a 3-6 fold higher GTPase activity than FtsZBs due to quicker disassembly of the FtsZEc polymers. The difference in GTP hydrolysis activity between FtsZBs and FtsZEc was reduced at 300 mM KCl, possibly because of reduced bundling of FtsZBs filaments (Figure 5).
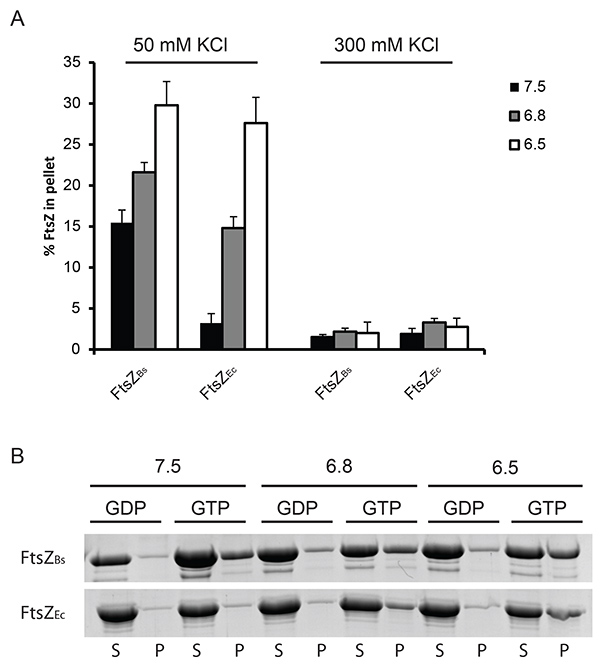
Figure 1. Quantification of FtsZ polymerization by sedimentation. (A) 12 µM FtsZ was polymerized in the presence of 2 mM GTP or GDP at pH 7.5 (black bars), 6.8 (grey bars) or 6.5 (white bars). The amount of protein pelleted was determined by densitometric analysis of Coomassie stained gels. GDP served as a control for a specific sedimentation and the percentage of FtsZ sedimented with GDP was subtracted from the percentage of FtsZ sedimented with GTP to obtain the values plotted in the graph. On the left: FtsZ sedimented at 50 mM KCl, on the right: FtsZ sedimented at 300 mM KCl. (B) Representative results from Coomassie stained gels. Polymerization of FtsZBs (upper gel) and FtsZEc (lower gel) at 50 mM KCl. (S) supernatant, (P) pellet fractions from the experiment. Click here to view larger image.
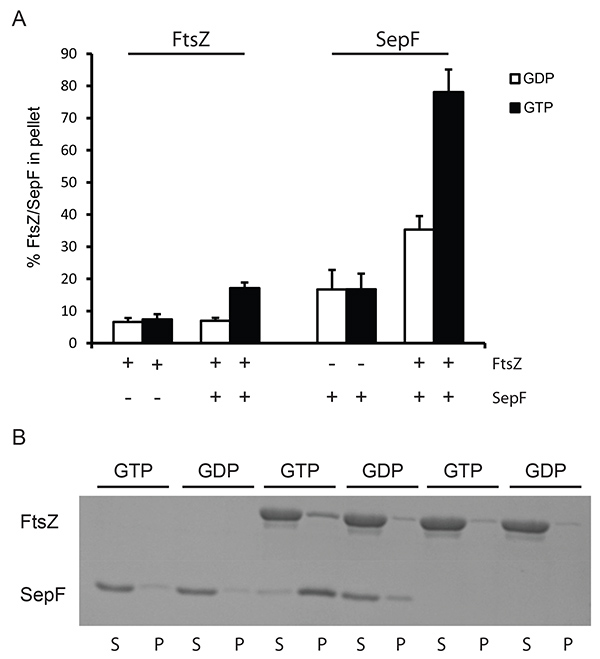
Figure 2. Sedimentation of SepF/FtsZ tubules at low speed. (A) 12 µM FtsZBs was polymerized with 2 mM GDP (white bars) or GTP (black bars). The amount of protein pelleted was determined by densitometric analysis of Coomassie stained gels. The + and – signs under the x-axis indicate the presence or absence of FtsZ and SepF in the reaction. (B) Representative results from a Coomassie stained gel. Polymerization of FtsZBs in the presence and absence of SepF. As a control SepF without FtsZ was used. Polymerization was carried out with GTP and GDP. (S) supernatant, (P) pellet fractions from the experiment. Click here to view larger image.
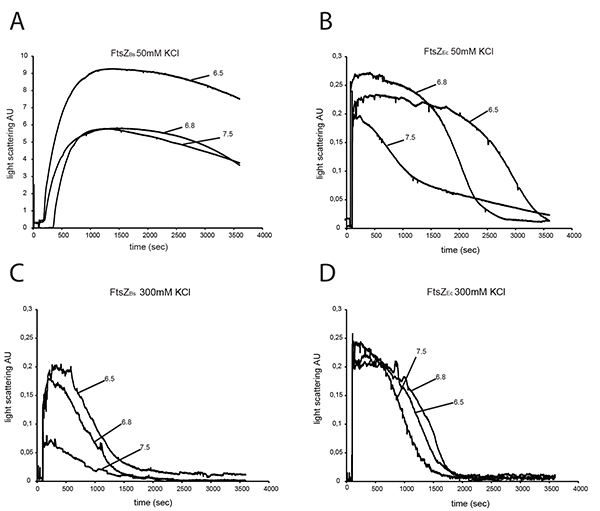
Figure 3. Light scattering of 12 μM FtsZEc and 12 μM FtsZBs. FtsZs were assembled in the presence of 2 mM GTP and polymerization was monitored by 90° angle light scattering. Polymerization of FtsZBs (A) and FtsZEc (B) at 50 mM KCl at pH 7.5, pH 6.8, pH 6.5. Polymerization of FtsZBs (C) and FtsZEc (D) at 300 mM KCl at pH 7.5, pH 6.8 and pH 6.5. Click here to view larger image.
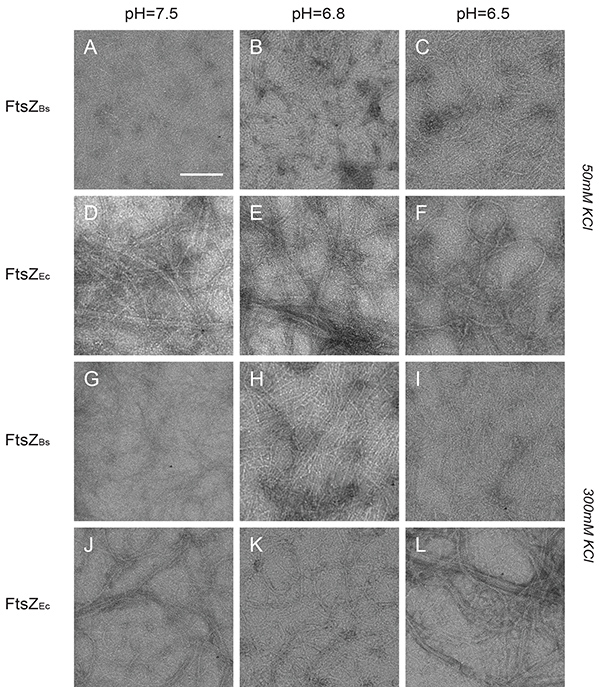
Figure 4. Structures of FtsZBs and FtsZEc polymers visualized by electron microscopy. (A-L) Images of 12 µM FtsZBs (A-C and G-I) and FtsZEc (D-F) and (J-L) polymerized with 2 mM GTP. (A-C) FtsZBs in buffer with 50 mM KCl and pH 7.5, 6.8, and 6.5 respectively. (D-F) FtsZEc in buffer with 50 mM KCl and pH 7.5, 6.8, and 6.5 respectively. (G-I) FtsZBs in buffer with 300 mM KCl and pH 7.5, 6.8, and 6.5 respectively. (J-L) FtsZEc in buffer with 300 mM KCl and pH 7.5, 6.8, and 6.5 respectively. Scale bar: 100 nm. Click here to view larger image.
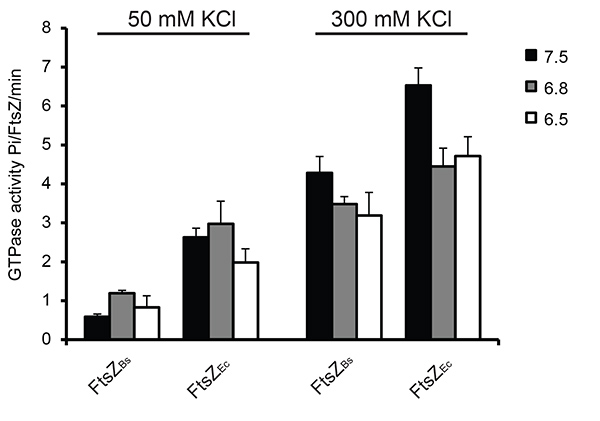
Figure 5. GTP hydrolysis during FtsZ polymerization in 6 different buffers. In all experiments 2 mM GTP was used. As a control sample with no MgCl2 was used. Activity of FtsZ without MgCl2 was subtracted from the activity of FtsZ in the presence of MgCl2. On the left: GTPase activity of FtsZ at 50 mM KCl, on the right: GTPase activity of FtsZ at 300 mM KCl. Click here to view larger image.
