As an example, we studied C. elegans in a quartz cuvette 1 cm wide, 5 mm thick and 4 cm tall cuvette. Sampling a single worm using video analysis, the average swimming frequency obtained from video analysis in a 5 mm thick cuvette is about 2.5 Hz (Figure 4). Similarly, sampling a single worm using the real time data acquisition method, we obtain a swimming frequency of about 2.7 Hz (Figure 5), using the digital oscilloscope (PicoScope).This procedure can be repeated for many worms. A detailed study of freely swimming worms revealed an average swimming frequency of 2.37 Hz in a 5 mm cuvette.6 As expected, the swimming frequency is higher than that for a crawling worm (~.8 Hz).3 Using this diffraction method, the average swimming frequencies of a C. elegans, which is confined to a microscope slide, has been found to match the previously published value of 2 Hz.1,7
Following procedures 3.) and then 6.) allows for the modeling of swimming diffraction patterns with the help of worm images obtained with a conventional microscope. The modeled diffraction patterns are used to simulate a swim cycle of the C. elegans (Figure 6). A successful model consists of physically feasible successive swim patterns matching the swimming frequencies. The worm should be in the same shape at the end of a swim cycle as it was in the beginning of a swim cycle.
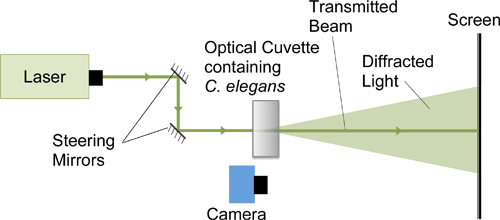
Figure 1. A green HeNe laser was used to create a dynamic diffraction pattern using live C. elegans. This diffraction pattern was filmed at 240 fps.
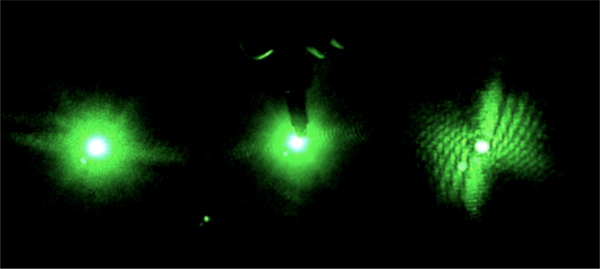
Figure 2. Drawing a black dot increases absorption of the transmitted beam. Saturation of the camera due to scattered light is reduced and the diffraction pattern becomes visible.
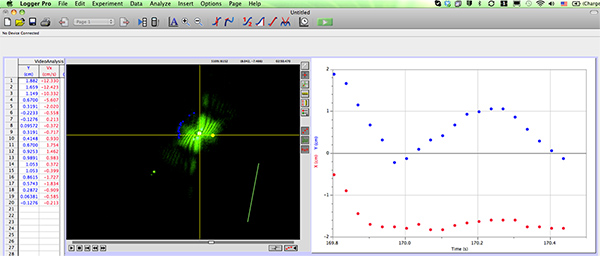
Figure 3. Screen shot of the video analysis software (Logger Pro) with a worm diffraction pattern that is being tracked. Click here to view larger figure.
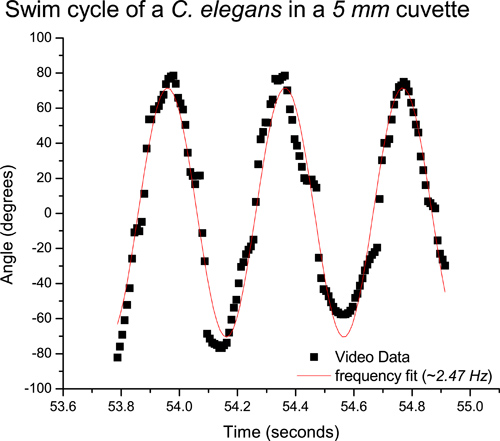
Figure 4. Video data corresponding to the swim cycle of a nematode in a 5 mm cuvette. The curve fit reveals a swimming frequency of ~ 2.5 Hz.
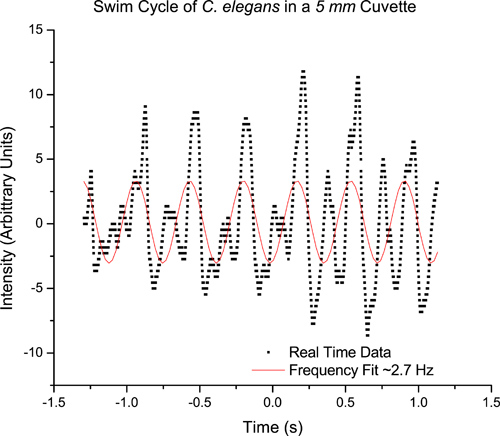
Figure 5. Real time data corresponding to the swim cycle of a nematode in a 5 mm cuvette. The curve fit reveals a swimming frequency of ~ 2.7 Hz.
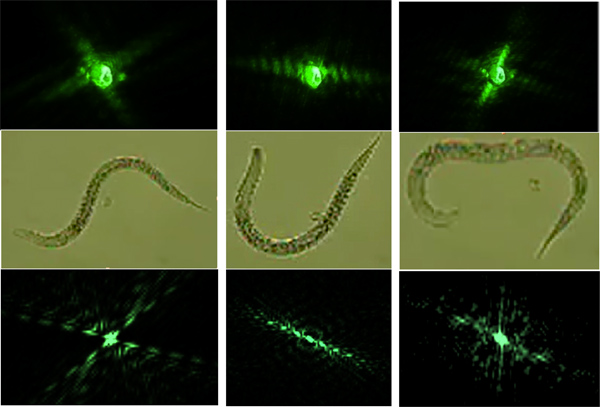
Figure 6. The top row represents the actual diffraction patterns and is matched to the modeled diffraction patterns in the bottom row. The modeled diffraction patterns were produced using worms on a microscope slide (middle row).
We have developed a novel approach to the real-time measurement of movement and simple locomotory behaviors in microscopic organisms like nematodes that does not require the use of microscopes.8 This methodological approach could also be utilized for studying numerous microscopic organisms like protists. This method is only limited by the wavelength of light used. The organism should not be smaller than the wavelength of the light. In addition to the cost-savings and portability of the equipment needed, one key advantage of this approach is the ability to measure behavior in real-time and in three dimensions, without the narrow constraints of image planes under a microscope. It is also possible with this technique to examine influences of gravitational forces or numerous other conditions on behavior that cannot be studied using microscope-based approaches.9 Thus, we can achieve a better understanding of microorganism natural locomotory behaviors freed from the confines of microscope slide droplets or specialized microfluidic chambers (Park et al, 2008).10
The lack of phase information in a diffraction pattern does not allow for the direct retrieval of the image corresponding to the diffracting object since the far-field diffraction pattern is proportional to the square of the absolute value of the Fourier transform. We are therefore calculating diffraction patterns from worm images so that they can be matched with the diffraction patterns of freely swimming nematodes (Figure 6).
This method has yielded results for truly freely swimming C. elegans and can be applied to any microscopic species that maneuvers in an optically transparent environment like water or many different ionic solutions. Conventional microscopes only allow studies with a focal depth on the order of micrometers.11 This is due to the limited depth of field when focusing light:
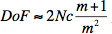
where the f-number N has a reciprocal relationship with the circle of confusion (c) so that a short focal length is associated with a large c.12,13 While this diffraction method is certainly not a replacement for conventional microscopy, it is able to deliver quantitative results quickly so that species can even be manipulated in real time at low cost. The diffraction patterns can be obtained with any laser pointer. The diffraction patterns can be filmed at a reduced temporal resolution using a regular digital camera. While the user may not have a microscope or a photodiode readily available, key parts of this experiment such as measuring thrashing frequencies and evaluating diffraction patterns can be completed at extremely low cost.
