All animal experiments described were approved by the University of Oklahoma or the University of California San Diego Institutional Animal Care and Use Committee. All steps handling infectious material were performed inside a biosafety cabinet (class II, type A2) and according to local regulations.
1. Tissue collection
- Infect appropriate trypanosomatid infection animal models, generally mice or hamsters.
NOTE: There is a considerable variety of mouse models for trypanosomatid infection, depending on the desired symptoms to be produced, speed of disease progression, disease severity, etc. The users can choose at their convenience.- For cutaneous leishmaniasis infection models, infect subcutaneously in the footpad or intradermally in the ear15.
- For visceral leishmaniasis infection models, infect intravenously1,17.
- For Chagas disease and sleeping sickness infection models, infect intraperitoneally6,12,18,19. Determine the parasite dose to use based on the parasite strain and planned time points.
- Plan sectioning positions.
- Plan to generate sections with a minimum of 10 mg of tissue per section. It is best to use 30-50 mg.
- For Chagas disease infection models, plan to collect cardiac and gastrointestinal segments systematically.
- For cutaneous leishmaniasis infection models, collect lesional tissue and lesion-adjacent samples.
- For visceral leishmaniasis infection models, plan to collect spleen and multiple liver lobes. Additional tissue sites such as adipose tissue may also be of interest to collect.
CAUTION: Samples from infected animals must be handled under the appropriate, institutionally-approved biosafety protocol. This will generally involve personal protective equipment (PPE) requirements and only opening tubes and collecting samples inside a biosafety cabinet (class II, type A2).
- Label and weigh tubes for homogenization.
- Use the tube type appropriate for the available homogenization system. For a TissueLyser, use 2 mL microcentrifuge tubes.
- Euthanize mice at the desired infection timepoints using isoflurane overdose as approved by institutional IACUC or according to IACUC-approved protocol.
- Section tissue systematically as planned, with one section per tube.
- Remember to wash sample collection equipment between samples with extraction solvent (50% methanol in this protocol).
- Keeping tube lid open, immediately snap freeze samples in liquid nitrogen.
CAUTION: Do not close tubes until any liquid nitrogen entered the tube has wholly evaporated to prevent tubes from exploding as nitrogen expands. Take adequate steps to ensure that the skin does not come in contact with liquid nitrogen (use cryo gloves and forceps to hold the tubes). Wear a safety face shield. - Store the tubes on dry ice until all the desired samples have been collected.
- Pause point: store samples at -80 °C until ready to do extractions.
- Weigh tubes to determine tissue sample weight. Record in a spreadsheet.
- Keep the samples frozen during the weighing process: keep tubes on dry ice, rapidly weigh, put back on dry ice right away. Do not allow the samples to thaw while weighing.
2. Metabolite extraction
NOTE: Only LC-MS grade liquids and reagents must be used throughout. This method was adapted from Reference20.
- Prepare all extraction solvents (LC-MS grade H2O, LC-MS grade methanol, spiked with 4 µM of sulfachlorpyridazine, LC-MS grade dichloromethane: methanol spiked with 2 µM of sulfachlorpyridazine) in 1 L glass bottles, using dedicated glassware.
NOTE: Volume to be prepared should be calculated based on sample weights, considering 500 µL of water per 50 mg of sample, 500 µL of methanol spiked with 4 µM of sulfachlorpyridazine per 50 mg of sample and 1,000 µL of prechilled dichloromethane: methanol spiked with 2 µM of sulfachlorpyridazine per 50 mg of sample, increasing the calculated volume by 10% to allow for pipetting inaccuracy. Store extraction solvent at 4 °C at least overnight to pre-chill. - Perform water-based homogenization of the tissue samples as per the steps mentioned below.
- Add one 5 mm stainless steel bead (see Table of Materials) to each of the 2 mL microcentrifuge tubes containing tissue samples, using a bead dispenser. Keep the tubes on ice.
- Make one blank tube containing LC-MS grade H2O that will go through all the steps and serve as an extraction blank. Use the average H2O volume from sample extractions. Add chilled LC-MS grade H2O to the frozen tissue samples.
- Normalize water volume to tissue weight by adding 500 µL of water / 50 mg of sample, using the sample weights calculated at step 1.7.
CAUTION: If handling biohazardous samples, continue to follow the appropriate, institutionally-approved biosafety protocol. This will generally involve requirements for personal protective equipment (PPE) and opening tubes only inside the biosafety cabinet (class II, type A2).
- Normalize water volume to tissue weight by adding 500 µL of water / 50 mg of sample, using the sample weights calculated at step 1.7.
- Homogenize samples at 25 Hz speed for 3 min using a tissue homogenizer (see Table of Materials).
- Close tubes tightly to avoid spilling the reagents during processing.
- Collect about 1/10th of the homogenization volume for DNA extraction, qPCR, protein-based analyses, or other analyses (if desired). Store in a microcentrifuge tube or 96-well plate (depending on the volume collected) for up to 6 months at -80 °C, if DNA extraction experiments are to be performed on a different day. Longer storage durations may be possible but have not been tested.
- Perform DNA extractions using any standard commercial kit for mammalian DNA extraction (see Table of Materials) from tissues as described in Reference13. Quantify DNA yield and store the extracted DNA at -20 °C. Acceptable DNA quantity and quality for qPCR has been observed even up to 3 years later.
NOTE: This step can be performed on frozen homogenate on a subsequent day from metabolite extraction. - Perform qPCR as described in Reference13, using 180 ng of extracted DNA.
NOTE: This can be performed on DNA collected on a previous day and frozen at -20 °C.- In the case of studies on T. cruzi infection, use the following primers: ASTCGGCTGATCGTTTTCGA and AATTCCTCCAAGCAGCGGATA to quantify parasite levels21 and the following primers to normalize to host DNA levels: TCCCTCTCATCAGTTCTATGGCCCA and CAGCAAGCATCTATGCACTTAGACCCC22 (see Table of Materials).
NOTE: The recommended qPCR cycles are as follows: denature at 95 °C for 10 min; perform 40 cycles at 95 °C for 30 s, and then 58 °C for 60 s, and finally 72 °C for 60 s. Perform melting curve analysis as appropriate for the available thermocycler. Process data using the ΔΔCt method23 to obtain relative parasite load between sampling sites. Absolute quantification can be obtained by comparing sample-derived ΔΔCt values to a standard curve generated from known amounts of parasites, spiked into uninfected tissue samples, and extracted as in steps 2.2 to 2.2.4.1.
- In the case of studies on T. cruzi infection, use the following primers: ASTCGGCTGATCGTTTTCGA and AATTCCTCCAAGCAGCGGATA to quantify parasite levels21 and the following primers to normalize to host DNA levels: TCCCTCTCATCAGTTCTATGGCCCA and CAGCAAGCATCTATGCACTTAGACCCC22 (see Table of Materials).
- Perform protein-based characterization of immune responses using multiplexed cytokine kits or standard commercial ELISA kits (see Table of Materials) as described in Reference13 on the stored homogenate.
- Perform DNA extractions using any standard commercial kit for mammalian DNA extraction (see Table of Materials) from tissues as described in Reference13. Quantify DNA yield and store the extracted DNA at -20 °C. Acceptable DNA quantity and quality for qPCR has been observed even up to 3 years later.
- Save at least half of the 500 µL of the homogenization volume for metabolite extraction.
- Perform aqueous metabolite extraction.
NOTE: Solvent selection can be adapted based on the chemical properties of metabolites of interest.- Add ice-cold LC-MS grade methanol spiked with 4 µM of sulfachlorpyridazine to the homogenate to achieve a final concentration of 50% methanol with 2 µM of sulfachlorpyridazine in water.
CAUTION: Methanol is flammable and hazardous. Use appropriate safety procedures, including handling inside a fume hood or biosafety cabinet. - Homogenize samples in a tissue homogenizer at 25 Hz speed for 3 min. Centrifuge samples at 16,000 x g at 4 °C for 10 min.
- Collect an equal volume of supernatant into a 96-well-plate. Select the volume to match the smallest volume of methanol + water combined across all samples. This is the aqueous fraction.
- Set aside any remaining aqueous homogenate supernatant at -80 °C as a backup.
- Keep the solid residue on ice while collecting the supernatants.
- Dry aqueous extraction supernatant until dry (~3 h or overnight). Use maximum speed and no heating.
- Freeze the dried 96-well-plate at -80 °C.
- Add ice-cold LC-MS grade methanol spiked with 4 µM of sulfachlorpyridazine to the homogenate to achieve a final concentration of 50% methanol with 2 µM of sulfachlorpyridazine in water.
- Perform organic metabolite extraction.
NOTE: Solvent selection can be adapted based on the chemical properties of metabolites of interest.- Add 1,000 µL per 50 mg of the sample of prechilled dichloromethane: methanol spiked with 2 µM of sulfachlorpyridazine to the solid residue from step 2.3.4.
CAUTION: Use appropriate safety procedures when handling solvents, including handling inside a fume hood with a good flow rate. - Homogenize samples in a tissue homogenizer at 25 Hz speed for 5 min. Centrifuge the samples at 16,000 x g at 4 °C for 10 min.
- Collect an equal volume of supernatant into a 96-well-plate. Select the volume to match the smallest volume of dichloromethane: methanol, across all samples. This is the organic fraction.
- Store the pellet at -80 °C as a backup. Store the remaining organic extract at -80 °C as a backup. Air-dry the organic extract in a fume hood overnight.
- Freeze the dried 96-well-plate at -80 °C.
- Add 1,000 µL per 50 mg of the sample of prechilled dichloromethane: methanol spiked with 2 µM of sulfachlorpyridazine to the solid residue from step 2.3.4.
3. LC-MS data acquisition
- Resuspend aqueous and organic extracts into 60 µL each of 50% methanol + 2 µM of sulfadimethoxine, and combine. Sonicate for 10 min; then, centrifuge for 10 min and transfer the supernatant to a clean 96-wellplate. Seal with zone-free plate seal and place the plate in an LC autosampler.
CAUTION: Use appropriate safety procedures when handling solvents, including handling inside a fume hood with a good flow rate. - Connect appropriate mobile phases to the LC system (see Table of Materials).
NOTE: For positive mode reversed-phase LC, authors recommend LC-MS-grade H2O + 0.1% formic acid as mobile phase A and LC-MS-grade acetonitrile + 0.1% formic acid as mobile phase B with a flow rate of 0.5 mL/min and a 7.5 min LC gradient. Recommended gradient steps are as published in Reference6: 0-1 min, 2% B; 1-2.5 min, linear increase to 98% B; 2.5-4.5 min, hold at 98% B; 4.5-5.5 min, linear decrease to 2% B; 5.5-7.5 min, hold at 2% B. - Ensure that the instrument is clean. Calibrate MS in both positive and negative mode.
- Perform MS performance evaluation as appropriate for the instrument.
- Create MS run sequence.
- Start with 2 blanks, 2 standards (6-mixes), and 5 pooled quality controls (QC) in a dilution series, beginning at 2 µL of injection volume and increasing stepwise to 30 µL injection volume.
- Randomize the sample order.
- After every 12 samples, run a blank, and then a pooled QC.
- Connect C8 LC column (1.7 µm particle size, 100 Å pore size, 50 x 2.1 mm length x internal diameter) and monitor for leaks and excessive backpressure. Fix issues as per instrument standard operating procedure.
- Start MS run sequence and collect data-dependent LC-MS/MS data.
NOTE: For a Q-Exactive Plus MS instrument, use Heated Electrospray Ionization and data-dependent MS2 acquisition (top 5) in positive mode, a resolution of 70,000 for MS1 and 17,500 for MS2, AGC Target of 1E6 for MS1 and 2E5 for MS2, maximum IT of 100 ms for both MS1 and MS2, scan range to 100-1500 m/z for MS1, and MS2 isolation window of 1 m/z. Set sheath gas to 35, aux gas to 10 and sweep gas to 0, spray voltage to 3.8 kV, capillary temperature to 320 °C, S-lens RF level to 50, and aux gas temperature to 350 °C.- Verify the data quality: check initial blanks (confirm lack of major peaks), standards (confirm the presence of expected peaks and symmetric peak shape), and QCs (confirm the presence of expected peaks, peak shape, and expected peak intensity).
- Periodically monitor MS run during the run sequence.
- Once the run is finished, check the data for any missed injections or other errors.
- Store the LC column as recommended by the manufacturer. Remove and store samples at -80 °C.
- Upload raw data to the data repository.
NOTE: MassIVE (massive.ucsd.edu) is recommended to enable the downstream link to molecular networking for metabolite annotations24,25.
4. LC-MS data processing
- Convert raw files to open format (.mzXML or .mzML) using MSConvert26.
- Upload raw data and mzXML or mzML data to the data repository.
- Generate feature table. There are multiple tools to do so (MZmine, MS-DIAL, openMS, XCMS, etc.27,28,29,30).
NOTE: MZmine is recommended because it is free, open-source, and can directly import mzXML files after MSconvert, has graphical user interface options for processing data and monitoring the impact of parameter selection, and can directly export to GNPS for molecular networking24.
NOTE: Follow tool documentation and use parameters appropriate for the available instrument. Additional details can also be found in Reference31.- Export feature table in .csv format.
5. 3D model generation
- Hand-draw 3D model de novo to scale as per the steps mentioned below.
- Take a picture of the organ of interest.
- Perform the following in SketchUp software (see Table of Materials) as mentioned below.
- Delete the default picture of a man that appears when the software opens.
- Click on File > Import to import a picture of the organs of interest.
- Click on the Lines tool and select the Freehand option. Use the pencil tool to trace and draw the outlines of the organs of interest. Ensure to close the line by drawing all the way back to the starting point. Once the line has been successfully closed, the drawn area will automatically appear shaded.
- Select the Push/Pull tool and pull up on the shaded area to convert the drawing from 2D to 3D.
- Delete the organ picture: select the Eraser button, and then right-click on the picture and select Erase.
- Export the file in .dae format: File > Export > 3D Model.
- Improve the realism of the model as per the steps mentioned below.
- Import the model into MeshLab software (see Table of Materials): open MeshLab and select File > Import Mesh. Select the .dae model generated at the previous step. A Pre-Open Options menu will pop up. Select OK.
- Select Wireframe on the top menu. Then, select: Filters > Remeshing, Simplification and Reconstruction > Subdivision surfaces: Midpoint. Leave all values as default and select Apply twice. Visually inspect the wireframe view of the model to ensure it is finely gridded. Close the pop-up menu.
- Export the model in .stl format: File > Export Mesh As, and then select STL File Format (*.stl) in the Files of Type dropdown menu. Click on Save. Select OK in the next pop-up menu.
- Open the Meshmixer software (see Table of Materials). Click on the Import (+) button. Select the .stl file generated at the previous step. Use the Sculpt > Brushes > Drag and Sculpt > Brushes > Inflate tools to pull out the model's surfaces that need to be rounded out.
- Once the model has the desired appearance, save it in .stl format: File > Export. Name the file as desired and select STL Binary Format (*.stl) in the Save as type dropdown menu. Click on Save. If a pop-up menu appears, click on Weitermachen.
6. 'ili plot generation
- Obtain coordinates for the positions in the 3D model that correspond to the sampling sites.
- Open the 3D model from step 5.1.3.5 in the MeshLab software: File > Import Mesh. Select the model generated in step 5.1.3.5. Click on OK in the Post-Open Processing pop-up window.
- To obtain x, y, and z coordinates for each sampling spot: select the PickPoints tool, and then right-click at regularly spaced intervals across the 3D model surface. Once all the desired coordinates have been selected, click on the top-most Save button in the Form pop-up window.
NOTE: This will export the coordinates in .pp file format. This file can be opened in spreadsheet software. - In spreadsheet software: Open the .pp file generated in step 6.1.2. Adjust data display using Data > Text to Columns > Delimited. Click on Next, and then select Space, and click on Finish.
- Reformat so that only numerical values remain in the spreadsheet cells by selecting Home > Find & Select > Replace. In the Find what box, enter: y=". Leave the Replace with box empty. Click on Replace All, and then on OK. Repeat for x=" and for z=" and for " />. Values are now ready for step 6.2.
- Make the 'ili feature table. This method was adapted from Reference16.
- In a spreadsheet software, build the feature table. Rows correspond to each position and columns to data.
NOTE: The first columns must be the position name (sample name), followed by x, y, and z coordinates of the sampling spots obtained at step 6.1.2 (with column headers x, y, z). The fifth column must be titled "radius". - Paste the appropriate metadata and metabolite feature abundance in the subsequent spreadsheet columns.
- In column "radius", enter the desired size of the sampling spots to be visualized on the model. Determine the values for radius empirically: enter 1 as default, and then assess whether radius or coordinates need to be adjusted in step 6.3. Save the file in .csv format (File > Save As), and then select CSV (Comma delimited) (*.csv) in the dropdown menu. Name the file as desired. Click on Save.
- In a spreadsheet software, build the feature table. Rows correspond to each position and columns to data.
- Open the data in 'ili (software developed by16).
- Open the 'ili website (ili.embl.de). Select Surface. Drag and drop the created 3D model into the browser window. Drag and drop the created feature table into the same browser window.
- Use the legend at the bottom-right corner to project the desired data column on the 3D model. Ensure that the spots and radii selected in step 6.2.1 match the sampling sites. If necessary, adjust values in the feature table, or choose additional coordinates in the MeshLab (see step 6.1.2).
NOTE: Visualization can also be improved by selecting the Spots > Border Opacity and setting the slider to its maximal value. - Consecutively select each data column to assess the distribution of this metabolite feature on the 3D model.
NOTE: This approach can also be used to visualize only specific metabolite features of interest, for example, those with p < 0.05 or a certain fold change between infected and uninfected samples, as determined in external statistical analysis tools. Features to visualize may also be selected outside of 'ili through machine learning approaches such as a random forest.
- Perform linear/log data visualization as per the steps mentioned below.
- Using the Mapping tab at the top right of the page, select Linear or Logarithmic in the Scale drop-down menu.
- Set the same scale for all plots if visualizing multiple features.
NOTE: The website automatically chooses the minimum and maximum for each data column to display. - To set the scale manually, deselect the Auto Min/Max option and enter the desired scaling. All data will now be displayed within the same scale.
- Change the color scale of the data (grayscale, blue-white-red, etc.).
- Change the color scale to the desired color scheme in the Mapping menu using the Color Map drop-down menu. Ensure that the selected color scheme is color-blind-friendly.
- To change the color of the 3D model or the background color, use Color and Background options under the 3D menu.
- Hide 3D axes by unselecting the Show the Origin box under the 3D menu.
- Save the image by screenshotting, using the snipping tool, or the shortcut key Crtl + S for Windows or Linux and
 + S on OS X.
+ S on OS X.
The number of metabolite features obtained depends on the tissue type analyzed and data processing parameters. For example, this protocol has been used to analyze the spatial impact of T. cruzi infection on the gastrointestinal tract metabolome in a mouse model of T. cruzi infection. In prior work, male C3H/HeJ were injected intraperitoneally with 1,000 CL + luc T. cruzi parasites32,6. Animals were euthanized 12 or 89 days post-infection, and a chemical cartography analysis of 13 contiguous segments of the gastrointestinal tract was performed as described in this protocol. This analysis led to a feature table of 5,502 features, which were then visualized into 3D using the steps described in this protocol. This approach enables the visualization of metabolite features in individual animals that are high at the site of high parasite load (kynurenine, Figure 2B vs. parasite load, Figure 2A), of metabolites with differential distribution across tissue regions (glutamine, Figure 2C) and metabolite features that are found at comparable levels across small and large intestines (LPE 16:0 Figure 2D). Kynurenine was selected for visualization because of its known relationship to inflammation and prior publications on the ability of kynurenine-derived metabolites to regulate T. cruzi load33. Random forest-based machine learning models had previously revealed an association between kynurenine levels and infection status6. Glutamine was selected for a visualization based on previous publications demonstrating a relationship between in vitro glutamine availability and T. cruzi drug sensitivity34. Differential distribution was confirmed using logistic regression, p < 0.05. LPE 16:0 was selected after visual inspection of the data to discover metabolite features found at comparable levels across tissue sites.
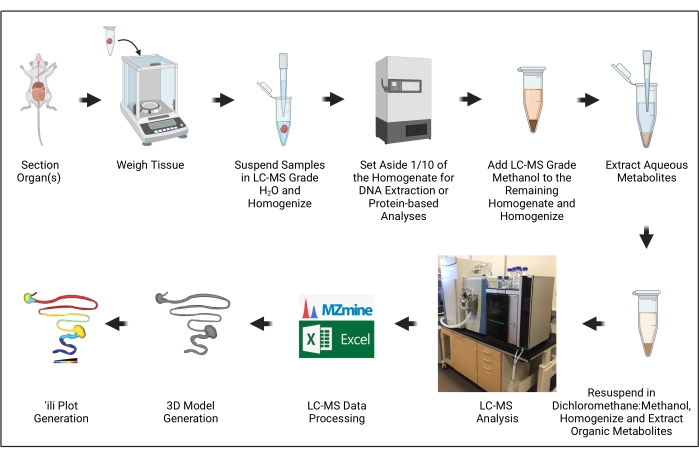
Figure 1: Protocol overview. The illustration was created with BioRender.com. Please click here to view a larger version of this figure.
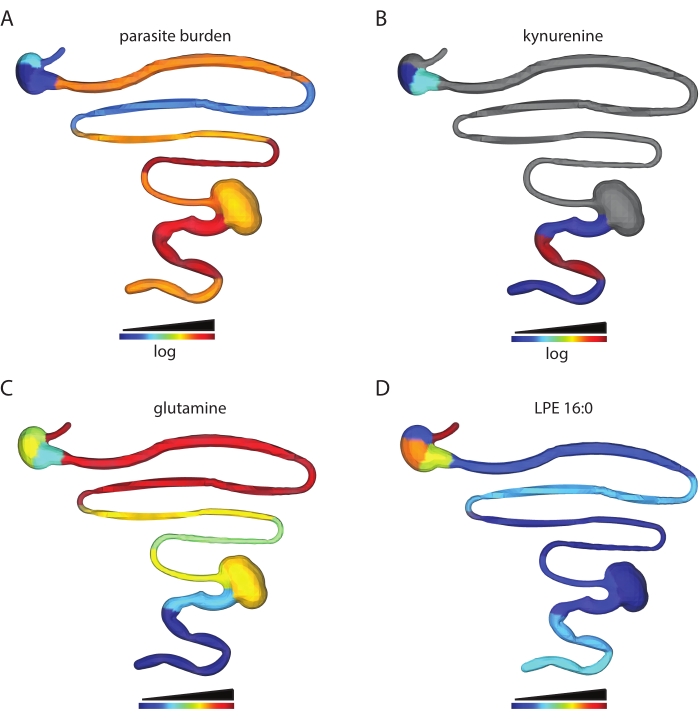
Figure 2: Chemical cartography analysis. Male C3H/HeJ mice were injected intraperitoneally with 1,000 CL+luc T. cruzi parasites32. Animals were euthanized 12 or 89 days post-infection, and the gastrointestinal tract was collected and sectioned systematically (step 1)6. Metabolites were extracted as in step 2 and analyzed by LC-MS/MS. 3D model generation was performed using the SketchUp software (step 5), and data were plotted in 3D as in step 6. (A) Parasite distribution in a specific mouse, 12 days post-infection. (B) Kynurenine metabolite distribution in the same mouse, 12 days post-infection. (C) Mean glutamine distribution across infected mice, 89 days post-infection. (D) Comparable levels of m/z 454.292 retention time 2.929 min, annotated as 2-hexadecanoyl-sn-glycero-3-phosphoethanolamine (LPE 16:0), in the same mouse as in A and B in the small intestine and colon. Samples and data were generated in6. Please click here to view a larger version of this figure.
