All animal handling and procedures were approved by the University of California, Riverside institutional animal care and use committee and were in accordance with guidelines from the American Association for Laboratory Animal Care, the United States Department of Agriculture, and the National Institutes of Health. The described method utilized C57BL/6 adult, female mice. All animals were euthanized by decapitation prior to tissue harvesting.
NOTE: An overview of the protocol, which uses a blue dye to assist in efficient dissection and uncoiling of the oviduct, is shown in the first figure (Figure 1).
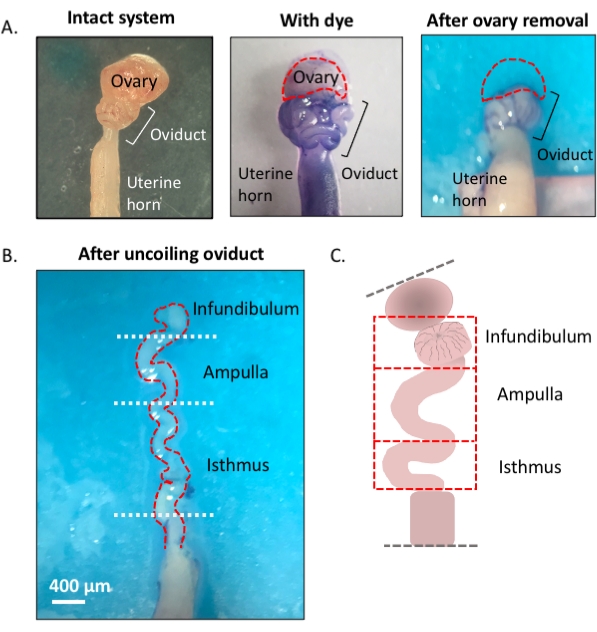
Figure 1: Overview of the microdissection method. (A) Left panel shows the intact structure from which most of the ovarian fat pad and remnants of connective tissue surrounding the oviduct have been removed. After this, the addition of toluidine blue helps distinguish the coils of the oviduct (middle panel). Right panel shows structures after the removal of the ovary. (B) An uncoiled oviduct with internal turns for determining where each segment begins and ends. (C) Cartoon of the segmentation used (not drawn to scale). Please click here to view a larger version of this figure.
1. Preparation of the dissection surface
- Affix a piece of dental wax to a Petri dish with adhesive and let it dry (Figure 2A). Sanitize the dish with 70% ethanol. The surface is ready for dissection.
2. Macrodissection of the ovary, oviduct, and uterus
- Immobilize the Petri dish containing the dental wax on a cold platform of choice (e.g., an ice pack kept at -20 °C prior to use) under a dissection microscope when ready for dissection (Figure 2A).
- Disinfect the ventral surface of the animal with 70% ethanol. Open the abdominal and pelvic cavity with sterile dissection scissors and move the gastrointestinal tract to one side to locate the dorsal bihornal uterus. Follow each uterine horn rostrally to locate each oviduct and ovary enveloped in the uterine fat pad, found just below the kidney (Figure 2B,C).
- Excise both lateral oviducts with the ovaries and a portion of the distal uterine horn still attached using dissection scissors. Cut each lateral uterine horn with approximately 1-2 cm of the horn still attached to the coiled oviduct and ovary (Figure 2D). Submerge the tissue immediately in 2 mL of cold dissection medium (see Table of materials).
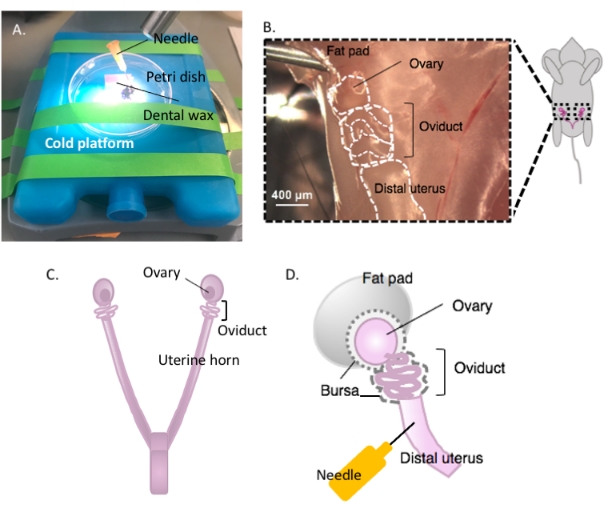
Figure 2: Macrodissection of the ovary and the dissection setup. The oviduct is located dorsally in the pelvic cavity at the base of the kidney within the ovarian fat pad (B). Locate the uterine bifurcation in the pelvic cavity (C) and follow the lateral uterine horn to locate the uterus-oviduct-ovary continuum (B). Remove these structures by cutting through one uterine horn and coarse dissection. Place onto the dissecting platform (A) and affix the uterine portion to dental wax with a needle (A and D). Please click here to view a larger version of this figure.
NOTE: Leave enough uterine horn intact and attached to the oviduct to allow affixation to the dissection platform (Figure 2D).
- Affix the uterine tissue to the dental wax with a sterile 25 G needle to secure tissue for dissection (Figure 2A,D, and Figure 3A). Working under the dissection microscope, clean and dissect away the ovarian fat pad and connective tissue to allow clear visualization of the oviduct (Figure 3B).
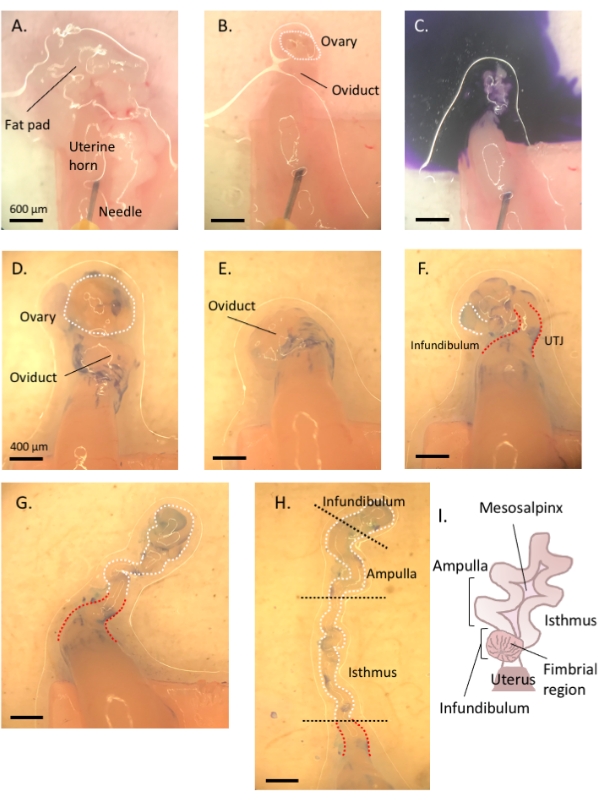
Figure 3: Step by step oviduct microdissection. Following removal of remnant fat and connective tissue (A,B), add toluidine blue to the tissue (C), and then wash away to facilitate visualization and subsequent removal of the ovary and uncoiling of the tube (D–H). The infundibulum can be found beside the proximal isthmus and uterine-tubal junction (UTJ). Lightly pull the distal end while cutting at the mesosalpinx to uncoil and reveal the endogenous turns of the oviduct (H) to orient for appropriate segmentation. Figure A-C scale bar = 500 μm, D–H scale bar = 400 μm (I) Cartoon of the various segments (not drawn to scale). Please click here to view a larger version of this figure.
- Using a 1 mL syringe, dispense and incubate the oviduct affixed to the uterus in 1-2 drops of sterile 1% toluidine blue dye solution for 30 s-1 min. Rinse with cold Dulbecco's phosphate-buffered saline (DPBS) and remove all the liquid (Figure 3C).
- Lightly pull the ovary from the coiled oviduct and cut away at the bursal membrane and broad ligament to remove the ovary from the oviduct without compromising the tubal tissue (Figure 3D,E).
NOTE: The ovary is not attached to the oviduct but encased in the bursal membrane with the oviduct (Figure 2D) and connected to the lateral side of the uterus via the broad ligament.
3. Microdissection and segmentation of the oviduct
- Locate the distal, fimbrial end of the oviduct found lateral to the uterine-tubal junction (UTJ). The oviduct coils back on itself; hence the fimbrial end can be located lateral to the proximal end and is an appropriate starting point to orient for uncoiling of the oviduct (Figure 3F,I).
- While gently pulling the tube, cut away at the mesosalpinx (Figure 3I) with spring-form micro-scissors to uncoil the oviduct (Figure 3G).
- Once uncoiled, cut the tube to produce the infundibular, ampullary, and isthmic regions (Figure 3H).
- Following excision of the infundibulum (fimbrial end and proximal stalk), excise the ampullary region by cutting after the second prominent turn of the tube (cut between turns two and three, approximately 1 cm in length). Finally, cut the remaining portion to the UTJ, which is the isthmic region (Figure 3H).
NOTE: The oviduct will not completely uncoil into a straight structure. The tube will maintain its turns (Figure 3H)3. Based on the subsequent turns following the ampullary region, one may further segment the remaining tube into the ampullary-isthmic junction and isthmus3. Following ampulla excision, cut between turns five and six to obtain the ampullary-isthmic junction; the remaining tube (turn six to the start of the UTJ) is the isthmus3.
- Following excision of the infundibulum (fimbrial end and proximal stalk), excise the ampullary region by cutting after the second prominent turn of the tube (cut between turns two and three, approximately 1 cm in length). Finally, cut the remaining portion to the UTJ, which is the isthmic region (Figure 3H).
- Immediately snap freeze dissected tissue segments in liquid nitrogen for RNA isolation or fix and process each segment for oriented embedding and immunohistological analysis.
- Add 200 μL of cold RNA extraction reagent and 1:1 homogenization bead mix (see Table of materials) to the tissue if proceeding for RNA extraction (see step 5.1).
4. Oviduct dissociation
- Continued from step 3.1.1 above) For optimal dissociation of the epithelium, slit open the oviduct in the areas where possible (i.e., infundibulum and ampulla) to expose the luminal epithelium (Figure 4A).
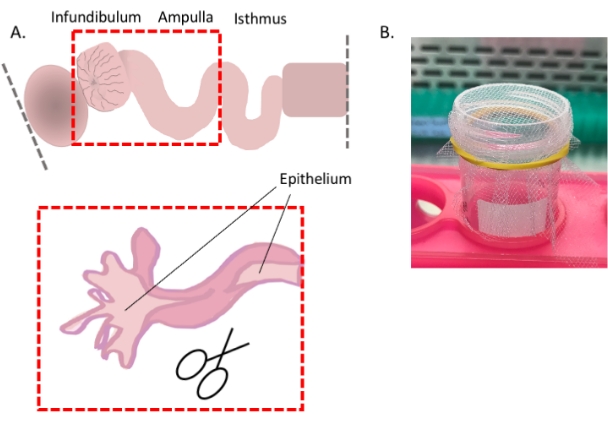
Figure 4: Oviduct cell dissociation, part 1. Cartoon showing how to expose the distal epithelium for optimal cell dissociation (A) and an image showing the easiest way to use the 1 mm2 mesh (B). Please click here to view a larger version of this figure.
- Starting at the fimbrial region, using forceps as leverage, slit the tube longitudinally with spring-form scissors.
- Mince the slit-open portions and the remaining oviduct into segments (~1-2 mm in size) and add 5 mL of warm, non-enzymatic dissociation buffer (see Table of Materials) and incubate at 37 °C.
- Resuspend the tissue pieces and dissociated cells with a bulb pipettor every 3 min for 9-10 min. Dilute the dissociation buffer 1:1 with cold dissection medium.
- Collect the cells by centrifugation (350 x g, 3 min, 4 °C). Resuspend the cells in 2 mL of RBC lysis buffer for 2 min at room temperature, and then dilute 1:1 with cold dissection medium.
- Collect the cells by centrifugation (350 x g, 3 min, 4 °C) and resuspend in 5 mL of cold pronase solution.
- Incubate in pronase digestion medium and resuspend the cell clumps with a bulb pipettor every 3-5 min until a single cell suspension is achieved (~25-30 min).
NOTE: This step is best performed in a tissue culture plate or flask to allow constant observation preventing unnecessary overexposure to pronase.
- Incubate in pronase digestion medium and resuspend the cell clumps with a bulb pipettor every 3-5 min until a single cell suspension is achieved (~25-30 min).
- Pass the cells through a sterile macro-mesh (1 mm x 1 mm mesh affixed to a conical tube with a rubber band (Figure 4B)) to remove any remaining undissociated tissue pieces. Collect the cells by centrifugation (350 x g, 3 min, 4 °C) and resuspend them in a cold medium appropriate for downstream analysis (e.g., FACS buffer if proceeding with staining for flow cytometry).
5. RNA extraction of oviductal segments
- Homogenize snap-frozen samples in 200 µL of RNA extraction reagent in a bead bullet blender tissue homogenizer on level 12 for two intervals (3 min each) at 4 °C using the stainless-steel bead mix.
NOTE: To preserve RNA integrity, samples should be immediately moved from snap-frozen state into RNA extraction reagent and not weighed. - Briefly centrifuge homogenate at 100-200 x g in a tabletop centrifuge for approximately 5 s at room temperature to separate the liquid from the beads. Transfer the supernatant to a fresh tube.
- Add another 200 µL of RNA extraction reagent to the beads to recover more samples, followed by centrifugation (100-200 x g, 5 s, room temperature). Combine the supernatants.
NOTE: Use a small pipette tip (most p200 tips are suitable) to prevent bead carryover when removing the supernatant. - Follow manufacturer's guidelines to phase separate and precipitate RNA. Representative samples were precipitated overnight at -20 °C with 100% isopropanol (approximately 2:1 volume, isopropanol: recovered aqueous phase).
- Transfer the complete RNA precipitate along with the solution to an RNA-binding spin column and centrifuge for 30 s at 9,500 x g. Purify the RNA on-column as per the manufacturer's guidelines, and then elute the RNA in 20 µL of nuclease-free water. Assess the quality/quantity on a microvolume spectrophotometer and/or bioanalyzer.
The described dissociation protocol yields 100,000-120,000 cells per mouse with pooling of both oviducts. The method is gentle enough to leave multi-ciliated cell borders intact, allowing for a distinction between multi-ciliated cells and secretory cells, and verifying that the digestion method is gentle enough to prevent de-differentiation. Representative immunofluorescence images in Figure 5 show small cell clumps following step 4.2.1, fixed for 3 min in 4% paraformaldehyde (PFA)/ DPBS, washed with 1% bovine serum albumin (BSA)/ Tris-buffered saline-tween (BTBST) and stained in suspension. Briefly, cells were permeabilized for 15 min at room temperature in 0.5% TritonX-100/BTBST, washed in BTBST, quenched for background fluorescence for 2 h at room temperature in 20 mM glycine/ DPBS, washed in BTBST, and then blocked for 1 h at room temperature in 5% BSA/ 0.1% TritonX-100/ TBST. Cells were then incubated in the primary antibody at 4 °C overnight in BTBST (mouse anti-mouse occludin (1:2000); rabbit anti-mouse acetylated tubulin (1:1000)) followed by several washes in BTBST. Secondary antibody was added for 1 h at room temperature in BTBST (goat anti-mouse IgG Alexa Fluor 555 (1:1000); goat anti-rabbit IgG Alexa Fluor 488 (1:1000)), washed several times in BTBST, once in DPBS, and then incubated in nuclear stain for 20 min at room temperature (1:2000, Hoescht 33342) followed by a wash in DPBS. Cells were resuspended in 80 µL of antifade mounting medium and left overnight in the dark before imaging (Figure 5A). Figure 5B shows cells at the end of the dissociation protocol. Viability assessments were made throughout the protocol, and representative images are shown in Figure 5B–D. Propidium iodide was added at 1:100 dilution, and cells were immediately observed on an inverted compound fluorescence microscope (Figure 5C, red stain). Cilia remained intact in small cell clusters (Figure 5A) and single cell suspensions (Figure 5E) with many cells observed to have cilia still beating (Suppl. Video 1).
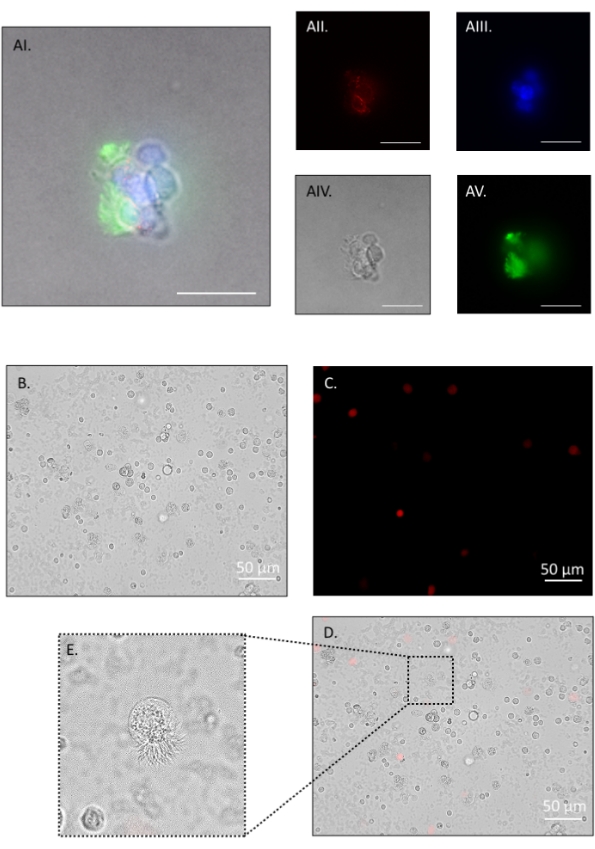
Figure 5: Oviduct cell dissociation, part 2. Fluorescent and phase-contrast images of a cell cluster positive for occludin (red), nuclei (blue), and acetylated tubulin (green). The ciliated apical border (green in AI and AV) remains intact throughout the protocol and withstands further digestion to single cells (E). AI: merge, AII: red channel, AIII: blue channel, AIV: phase contrast, AV: green channel. After a brief pronase incubation, the majority of cells are single. Propidium iodide (PI) staining shows approximately 93% viability. (B) Bright field, (C) red channel PI positive, (D) merge. (E) inset on merge shows an intact ciliated border on a single dissociated cell. Such cells had actively beating cilia (see Suppl. Video 1). Images were taken on an inverted compound fluorescence microscope. Figure AI-V scale bar = 20 μm, B-D scale bar = 50 μm. Please click here to view a larger version of this figure.
The RNA isolation protocol described gives sufficient yield and purity necessary for segmental RT-qPCR and RNAseq. Results show that this method yields 800-1200 ng RNA per segment (Figure 6A) except for infundibular samples where pooling from two animals may be necessary for appropriate yield for downstream assays. The presented results for infundibulum are pooled from two animals, totaling four infundibular regions (Figure 6). This protocol is successful in achieving pure RNA, initially analyzed by microvolume spectrophotometer and further by bioanalyzer (Figure 6B,C). Samples have a 260/280 nm absorption ratio between 1.8 and 2.0 (Figure 6B) typical of pure RNA nucleotide species19. Absorption ratios (260/230 nm) were taken for each sample with an inclusion criterion between 1 and 2.2, representative of limited contamination from isolation reagents (data not shown)19. Bioanalyzing the samples showed high integrity of the RNA, with assessed RNA integrity number (RIN) above seven (Figure 6C), appropriate for downstream expression and sequencing analysis20.

Figure 6: Quality assessment of segmental RNA. Whole RNA was assessed for yield (A) and quality (B,C) by microvolume spectrophotometer and bioanalyzer. RIN: RNA integrity number, as assessed by the bioanalyzer. Bars are the mean ± standard deviation (SD), N = 12 samples per segment from two separate RNA extractions. Please click here to view a larger version of this figure.
Supplemental Video 1: Live cell imaging of beating cilia on dissociated cells. Beating cilia following pronase digestion. Videos were taken on an inverted compound microscope. Please click here to download this Video.
