The principle of this method is simple: the perfusate flows from the left chamber, the aortic valve is opened, and the perfusate runs into the coronary artery in the same direction as the blood run, since the aorta is closed by clamping, which enables the deep perfusion of the myocardium in order to digest the extracellular matrix.
Ventricular myocytes freshly isolated with the present method are shown in Figure 1A. Figure 1B shows enlarged images of the ventricular and atrial myocytes. This isolation procedure resulted in a high yield (70%-80%) of rod-shaped quiescent ventricular myocytes from adult mice (8-10 weeks), which were available within roughly 5 h after isolation (Figure 1C), a similar interval to that when using the traditional Langendorff-based procedure7. However, the ratio of freshly isolated viable cells was lower in aged mice of >2 years old (Figure 1C). The total number of ventricular myocytes obtained per adult heart using this protocol was approximately 3 x 106 cells, which was similar to the value previously reported7,12. The action potentials recorded in the ventricular and atrial myocytes (Figure 1D) were similar to those in cells obtained by the Langendorff-based method10. An immunostaining analysis confirmed that the sarcomeric structure of the ventricular myocytes was well-organized with a clearly visible cell membrane (Figure 2A). The individual cardiomyocytes isolated with this method can be directly used in experiments, such as an electrophysiological analysis10 or immunostaining experiment.
Cardiac fibroblasts exist in interstitial spaces. Sufficient digestion of the extracellular matrix results in the isolation of those cells. The isolated cardiac fibroblasts proliferate under culture conditions and can be passaged several times or stored in liquid nitrogen in an appropriate cell reservoir solution. Figure 2B shows that most of the cultured cardiac fibroblasts had transformed into myofibroblasts during subculture, as confirmed by the increased expression of α-smooth muscle actin13,14. Also, the cardiac progenitors can be isolated with the present method and cultured in appropriate culture medium, which start beating automatically10,15.
Homogenization of the robust myocardium is not easy, especially for cardiac tissue from aged mice, which possesses a large amount of extracellular fibers. After antegrade perfusion, protein from the atria and ventricles can be easily homogenized in the lysis buffer with light force to extract proteins. A Western blot analysis demonstrated the specific expression of ANP in atria but not in ventricles from adult (20 weeks old) and aged (108 weeks old) mice (Figure 3).
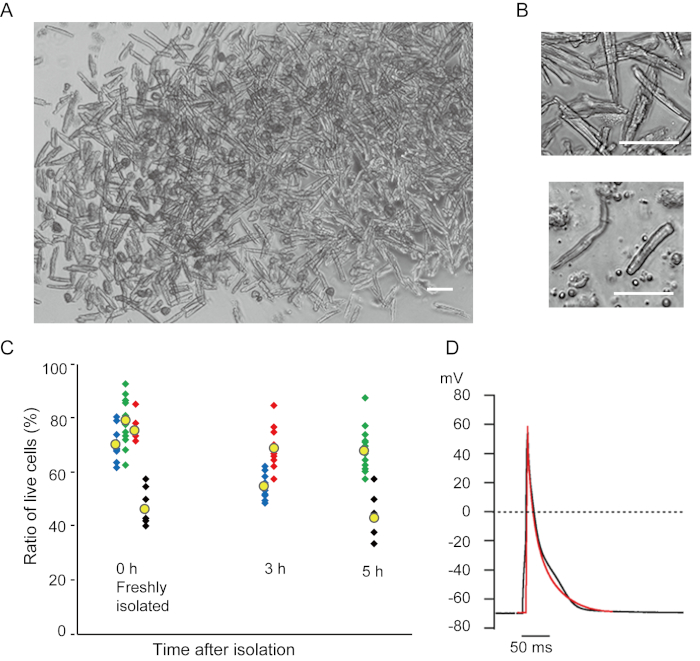
Figure 1. Isolated cardiomyocytes from mice. A. Ventricular myocytes freshly isolated with the antegrade perfusion, with images acquired with low magnification. After the final washing, the cardiomyocytes were resuspended with 2 mL of cell resuspension solution, 100 µL of which was dropped onto the glass-bottom culture dish and cell settlement awaited. Bar, 100 µm. B. Enlarged images of isolated ventricular myocytes (upper) and atrial myocytes (lower). Bar, 100 µm. C. Isolated cells were suspended in the cell resuspension solution and stored at 37°C for the desired period, and the number of live ventricular myocytes was counted in 10-15 fields under a microscope. Rounded cells were considered to have been irreversibly injured or dead16. Green, blue and red symbols were obtained from 3 mice at 8-10 weeks old, and black symbols were from 106 weeks old mouse. Yellow symbol indicates the mean of each group. D. Representative action potentials recorded from ventricular (black) and atrial (red) myocytes of 8-10 mice. The data were obtained from the cells approximately 3 h after isolation. Please click here to view a larger version of this figure.
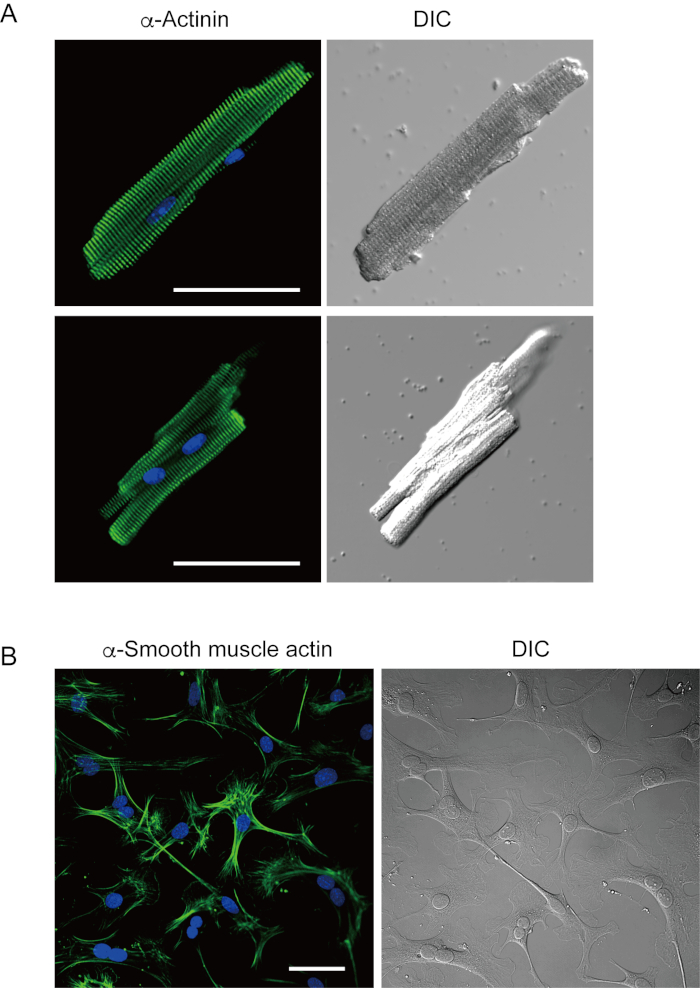
Figure 2. Immunostaining for α-actinin in isolated mouse ventricular myocytes and α-smooth muscle actin in cultured mouse cardiac fibroblasts. A. Confocal laser scanning microscopy of immunostaining for α-actinin (green), DAPI staining for nuclei (blue) and a DIC image of ventricular myocytes isolated from mouse heart with antegrade perfusion. Bar, 50 µm. B. Immunostaining for α-smooth muscle actin (green), DAPI staining for nuclei (blue) and a DIC image of cardiac fibroblasts isolated from mouse heart with antegrade perfusion. Cardiac fibroblasts were cultured for four days. Bar, 100 µm. Please click here to view a larger version of this figure.
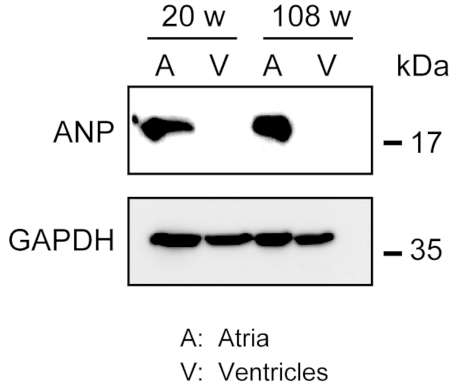
Figure 3. Western blot analyses of ANP in atria and ventricles. Western blot analyses for the atrial marker atrial natriuretic peptide (ANP) in atria (A) and ventricles (V) prepared from adult (20 weeks) and aged (108 weeks) hearts. ANP is present in the atria but absent in the ventricles. Use glyceraldehyde 3-phosphate dehydrogenase (GAPDH) as a control house-keeping protein. Please click here to view a larger version of this figure.
| Stock solutions for isolating heart cells | |
| 10X CIB (500 mL) | |
| NaCl | 37.99 g |
| KCI | 2.01 g |
| 1 M MgCl2 | 2.5 mL |
| NaH2PO4 | 0.23 g |
| HEPES | 29.79 g |
| DW | Fill up to 500 mL |
| 100 mM CaCl2 stock soution | |
| CaCl2 | 100 mM |
| 400 mM EGTA stock solution | |
| EGTA | 400 mM |
| Insulin solution | |
| Insulin | 1 unit/mL in 0.1 M HCl |
| 50X Antibiotics stock solution (20 mL) | |
| Penicillin | 100 mg |
| Streptomycin | 100 mg |
| Phenol red | 1.5 g |
| DW | 20 mL and sterilize with filtering |
| 10X Tyrode solution (1000 mL) | |
| NaCl | 81.82 g |
| KCl | 4.03 g |
| 1 M MgCl2 | 5 mL |
| NaH2PO4 | 0.47 g |
| HEPES | 11.92 g |
| NaOH | 0.8 g |
| DW | Fill up to 1000 mL |
| Stock solutin for immunostaining | |
| DAPI stock | |
| DAPI | 2 mg/mL in methanol |
| Stock solutions for Western blots | |
| HEPES buffer (100 mL) | |
| NaCl | 0.88 g |
| 400 mM EGTA | 0.25 mL |
| HEPES | 0.24 g |
| 1M NaOH | adjust pH to 7.4 |
| DW | Fill up to 500 mL |
| Protease inhibitors cocktail | |
| Complete mini | 1 tablet |
| DW | 0.4 mL |
Table 1. Description of the stock solutions. Keep stock solutions at 4 °C. Aliquot protease inhibitors cocktail for storage at -20 °C.
| Solutions for isoltaing heart cells | |
| CIB (200 mL) | |
| 10X CIB | 20 mL |
| Insulin solution | 0.01 mL |
| Glucose | 0.79 g |
| 1M NaOH | pH adjust to 7.4 |
| DW | Fill up to 200 mL |
| Enzyme-mix solution (30 mL) | |
| Collagenase type2 | 30 mg |
| Trypsin | 1.8 mg |
| Protease | 1.8 mg |
| 100 mM CaCl2 stock solution | 0.09 mL |
| CIB | 30 mL |
| CIB-Ca2+-BSA (15 mL) | |
| BSA | 30 mg |
| 100 mM CaCl2 stock solution | 0.18 mL |
| CIB | 15 mL |
| CIB-EGTA (150 mL) | |
| 400 mM EGTA stock solution | 0.150 mL |
| CIB | 150 mL |
| Tyrode solution (1000 mL) | |
| 10X Tyrode stock solution | 100 mL |
| Glucose | 0.99 g |
| 1M CaCl2 | 1.8 mL |
| 1M NaOH | pH adjust to 7.4 |
| DW | Fill up to 1000 mL |
| Cell resuspension solution (15 mL) | |
| BSA | 30 mg |
| 50X Antibiotics stock solution | 0.3 mL |
| Tyrode solution | 15 mL |
| Solutions for immunostaining | |
| Cell adherent solution (0.3 mL) | |
| Cell-Tak | 0.01 mL |
| 0.1 M NaHCO3 (pH8.0) | 0.285 mL |
| 0.1 M NaOH | 0.005 mL |
| Blocking-permeabilizatin solution (10 mL) | |
| Fetal bovine serum | 1 mL |
| Triton X-100 | 1 mL |
| 10X PBS | 1 mL |
| DW | 7 mL |
| K+ rich pipette solution | |
| Potassium aspartate | 70 mM |
| KCl | 50 mM |
| KH2PO4 | 10 mM |
| MgSO4 | 1 mM |
| ATP disodium salt | 3 mM |
| GTP lithium salt | 0.1 mM |
| EGTA | 5 mM |
| HEPES | 5 mM |
| KOH | pH adjust to 7.2 |
| Solutions for Western blots | |
| Lysis buffer (1 mL) | |
| HEPES buffer | 0.86 mL |
| Nonidet-P40 | 0.1 mL |
| Protease inhibitors cocktail | 0.04 mL |
| Runnning buffer (1000 mL) | |
| 10X TG (0.25 M Tris and 1.92 M Glycine) | 100 mL |
| SDS | 1 g |
| DW | 900 mL |
| Transfer buffer (1000 mL) | |
| 10X TG | 100 mL |
| Methanol | 200 mL |
| DW | 700 mL |
| Blotting buffer (TBST) (1000 mL) | |
| 5M NaCl | 20 mL |
| 2M Tris-HCl (pH 7.5) | 5 mL |
| 10% Tween 20 | 10 mL |
| DW | 965 mL |
Table 2. Description of the working solutions for isolating heart cells, immunostaining and Western blotting. Prepare all working solutions just before the experiments.
Supplementary Figure 1. Outline of the cell isolation. Flow diagram of the isolation of ventricular and atrial myocytes and cardiac fibroblasts from a single heart. Please click here to download this figure.
