Figure 1 shows representative brightfield images of several time points to demonstrate what the cells/organoids look like throughout the different stages of the protocol. The hESCs were removed from the tissue culture plate, broken into small pieces, and placed in a T75 ultra-low attachment flask where they formed spheres. It is important to note that the cells look bright and similar in size, without dark, dying cells in the centers of these clusters. The cells were gradually weaned off bFGF. On day 5, they were placed into neural induction media and they remained in this media throughout the culture period. Although the organoids get larger and thus darker over time, it is important to take note of the neural rosette-like structures (black arrows) that are present throughout the brain organoid development and expand. The rosettes indicate the initiation of neural differentiation and contain features of the embryonic neural tube, displaying epithelial characteristics and surrounding an apical lumen15.
Staining of the organoids with hematoxylin and eosin at 5 months in culture indicated that there were not vast amounts of necrosis even in the centers, which was of initial concern given the stagnant culture system (Figure 2A). These organoids demonstrated a histologic morphology similar to the human cortex based on light microscopic evaluation by an experienced neuropathologist (Figure 2B). By histology, many unique cell morphologies were observed resembling glia (blue arrow head), neurons (red arrowhead), cells with Cajal-Retzius morphology (black arrows), and neuropil (orange arrow head) (Figure 2B,C).
To take a more in depth look at gene expression within the cells, qRT-PCR was performed. For the results shown in Figure 3, each bar represents 3 separate batches of cells grown independently and harvested at the specified time point. These samples were then run in triplicate with a primer pair to the indicated gene in addition to the housekeeping gene, GAPDH. The glutamate transporter, Vglut1 (Figure 3A), was expressed at 2.5 weeks, increased at 5 weeks, and remained consistent through 5 months in culture. A forebrain marker, Foxg1 (Figure 3B), was expressed at low levels until 5 weeks in culture. The deep layer marker, Tbr1 (Figure 3C), peaked around 5 weeks and decreased subsequently, whereas the upper layer marker, Satb2 (Figure 3D), increased over time.
The expression of the ventral marker Engrailed1 (Eng1) (Figure 3E), the hindbrain/spinal cord marker Hoxb4 (Figure 3F), as well as the oligodendrocyte marker, Olig2 (Figure 3G), all increased over time. In contrast, the stem cell marker, Sox2 (Figure 3H), decreased over time. The glial marker, FAP (Figure 3I), peaked at 5 weeks and remained relatively constant subsequently. In addition, immunofluorescence data was consistent with the qRT-PCR data. At 10 weeks there was a robust expression of Foxg1 (Figure 4A). Sox2 expression was more confined to areas resembling the subventricular zone (SVZ) (Figure 4B,C). Interestingly, there was also some expression of the outer radial glial cell marker, HopX (Figure 4D).
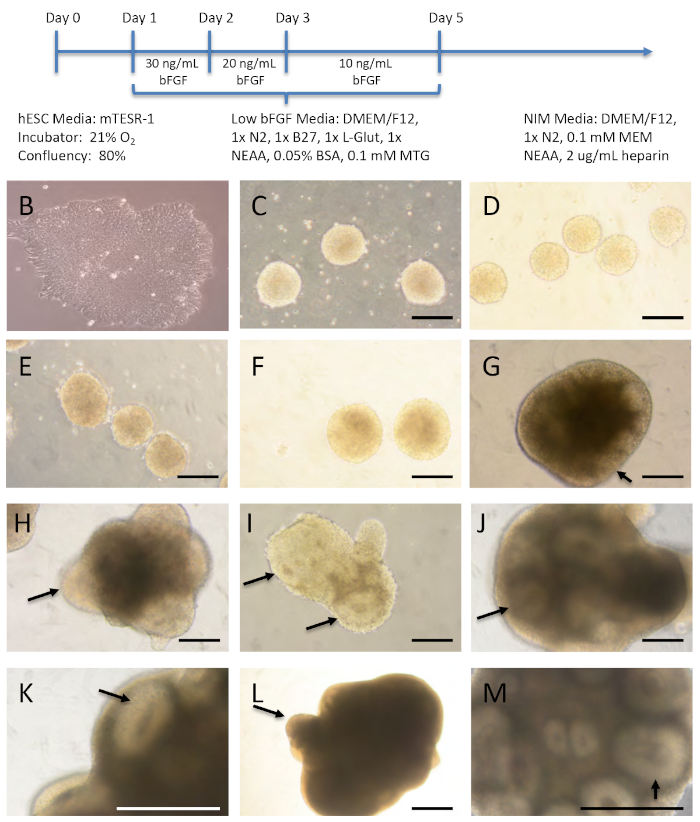
Figure 1: Overview of organoid growth conditions and morphology. (A) Schematic of media changes. (B-M) Representative images of organoids as they matured. (B-M) H9 hESCs (B) were utilized to form the brain organoids. Organoids on (C) day 2 in 20 ng/mL bFGF media, and (D) day 3 and (E) day 4 in 10 ng/mL bFGF media. (F-M) Organoids in neural induction media (NIM) on days 5 (F), 8 (G), 10 (H), 17 (I) 35 (J,K), and 70 (L,M). Arrows point to neural rosettes. Scale bar = 200 µm. Please click here to view a larger version of this figure.
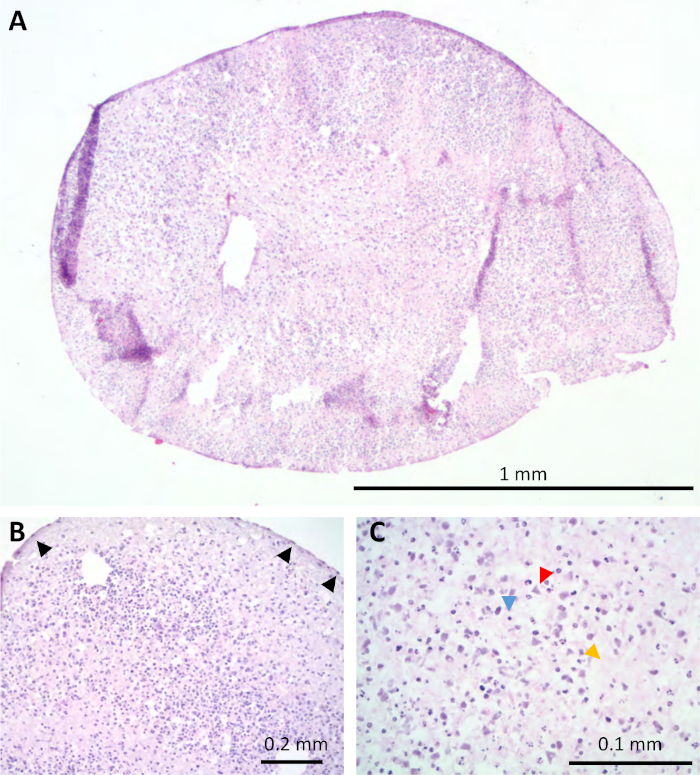
Figure 2: Organoids shared histologic similarities to human brain tissue. H&E staining of the organoids at 5 months (A) with some layering resembling the human cortex (B). At higher magnification many cell morphologies were observed including glia (blue arrow head), neurons (red arrowhead), neuropil (orange arrow head), and cells with Cajal-Retzius morphology (black arrows) (B,C). Please click here to view a larger version of this figure.
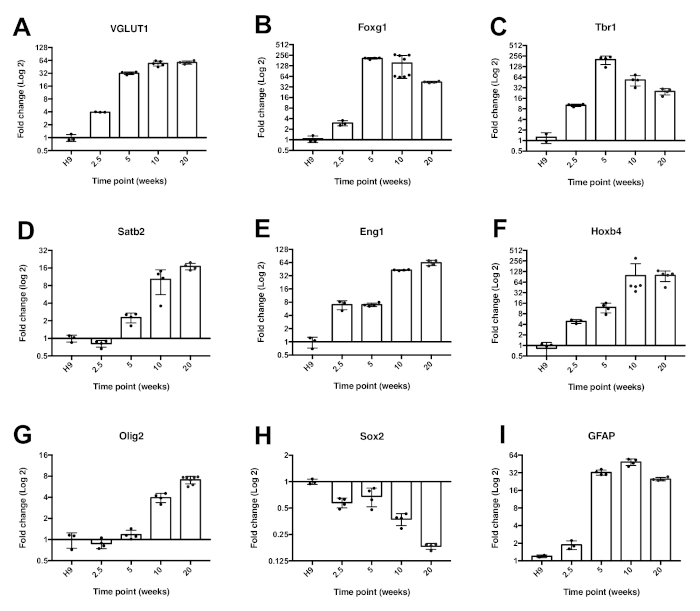
Figure 3: Expression of neurodevelopmental genes within the brain organoids over time. Quantitative RT-PCR data using SYBR green evaluating the expression of Vglut1 (A), Foxg1 (B), Tbr1 (C), Satb2 (D), En1 (E), Hoxb4 (F), Olig2 (G), Sox2 (H), and GFAP (I). Error bars = mean ± standard deviation (n ≥ 3). This figure has been modified from16. See Table 1 for primer information. Please click here to view a larger version of this figure.
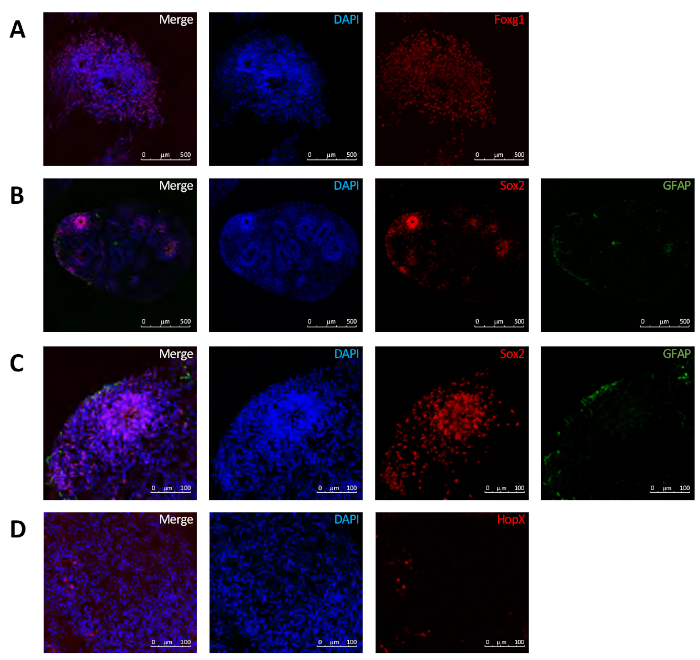
Figure 4: Expression of neurodevelopmental proteins within the brain organoids at 10 weeks. Immunofluorescence revealed a robust expression of Foxg1 (A), localized expression of Sox2 (B,C) and the presence of HopX (D). Please click here to view a larger version of this figure.
| Gene | Sequence (5' to 3') | Amplicon | Exon | |
| En1 | F | GGACAATGACGTTGAAA CGCAGCA |
149 | 2 |
| R | AAGGTCGTAAGCGGTTT GGCTAGA |
2 | ||
| Foxg1 | F | AGAAGAACGGCAAGTAC GAGA |
188 | 1 |
| R | TGTTGAGGGACAGATTG TGGC |
1 | ||
| GAPDH | F | ACCACAGTCCATGCCAT CAC |
449 | 8 |
| R | CACCACCCTGTTGCTGT AGCC |
9 | ||
| GFAP | F | AGAGATCCGCACGCAGT ATG |
80 | 4 |
| R | TCTGCAAACTTGGAGCG GTA |
5-Apr | ||
| Hoxb4 | F | AAAGCACCCTCTGACTG CCAGATA |
80 | 2 |
| R | ATGGGCACGAAAGATGA GGGAGA |
2 | ||
| Olig2 | F | CCCTGAGGCTTTTCGGA GCG |
451 | 1 |
| R | GCGGCTGTTGATCTTGA GACGC |
2 | ||
| Satb2 | F | TAGCCAAAGAATGCCCT CTC |
94 | 6 |
| R | AAACTCCTGGCACTTGG TTG |
7 | ||
| Sox2 | F | CCCAGCAGACTTCACAT GT |
150 | 1 |
| R | CCTCCCATTTCCCTCGT TTT |
1 | ||
| Tbr1 | F | GTCACCGCCTACCAGAA CAC |
101 | 4 |
| R | ACAGCCGGTGTAGATCG TG |
6 | ||
| Vglut1 | F | CAGAGTTTTCGGCTTTG CTATTG |
183 | 5-Apr |
| R | GCGACTCCGTTCTAAGG GTG |
6 |
Table 1: Primer sequences used for quantitative RT-PCR in Figure 3.
