The purpose of this protocol is to evaluate the effect of anti-c-fms antibody on osteoclast formation in the presence of RANKL or TNF-α, and to determine the effect of M-CSF on osteoclast precursors proliferation. In this protocol, we have provided a reliable process by which large quantities of pure osteoclast cultures are generated. We have also provided a way to test anti-c-fms antibody suitable concentration for inhibiting osteoclast formation under different culture conditions.
The osteoclast formation in M-CSF+RANKL culture with anti-c-fms antibody is shown in Figure 1. At 1, 10 ng/mL anti-c-fms antibody, there was no significant decrease in the number of osteoclasts compared to control (0 ng/mL). While at 100, 1,000 ng anti-c-fms antibody, there was a significant decrease in the number of osteoclasts compared to control. The osteoclast formation in M-CSF+TNF-α culture with anti-c-fms antibody is shown in Figure 2. At 1 ng/mL anti-c-fms antibody, there was no significant decrease in the number of osteoclasts compared to control (0 ng/mL). While at 10, 100, 1,000 ng/mL anti-c-fms antibody, there was a significant decrease in the number of osteoclasts compared to control.
Osteoclast precursors culture with M-CSF+anti-c-fms antibody is shown in Figure 3. At 1, 10, 100 ng/mL, there was no significant decrease in the number of osteoclast precursors compared to control (0 ng/mL), while at 1,000 ng/mL, there was a significant decrease in the number of osteoclast precursors compared to control. This indicates that a concentration as high as 1,000 ng/mL is needed to inhibit proliferation of osteoclast precursors significantly.
These results provide information about the robust nature of RANKL in producing osteoclasts while a concentration of 50 ng/mL of RANKL was used, and a concentration of 100 ng/mL of TNF-α was needed to obtain the same number of osteoclasts. Both of these methods are suitable for osteoclast generation but the decision of which is superior depends on the downstream application in the context of the reagents to be used later. As the results indicate, using RANKL is not the equivalent of using TNF-α to generate osteoclasts.
In this protocol, we used anti-c-fms antibody to inhibit osteoclastogenesis, and by looking at the results, we find that a concentration of 100 ng/mL of anti-c-fms antibody was needed to produce significant decrease in osteoclast number in RANKL wells. A concentration of 1,000 ng/mL of anti-c-fms antibody was needed to inhibit osteoclast precursor proliferation in MCSF proliferation assay, while a concentration of only 10 ng/mL of anti-c-fms antibody was needed to produce significant decrease in osteoclast number in TNF-α wells.

Figure 1: Effect of anti-c-fms antibody on RANKL-induced osteoclast formation. Microscopic observation of osteoclast precursors that were treated with M-CSF, RANKL and various dose of anti-c-fms antibody (0, 1, 10, 100 and 1,000 ng/mL) for 4 days. Cells were fixed and stained with TRAP staining. Number of TRAP-positive cells cultured with M-CSF, RANKL and various concentrations of anti-c-fms antibody (0, 1, 10, 100 and 1,000 ng/mL) for 4 days. Cells were fixed and stained with TRAP staining. The number of TRAP-positive cells was assessed. Results were expressed as mean ± S.D. of four cultures. Statistical differences were detected by using Scheffe's tests (n = 4; *p < 0.01). Scale bars = 200 µm. Please click here to view a larger version of this figure.
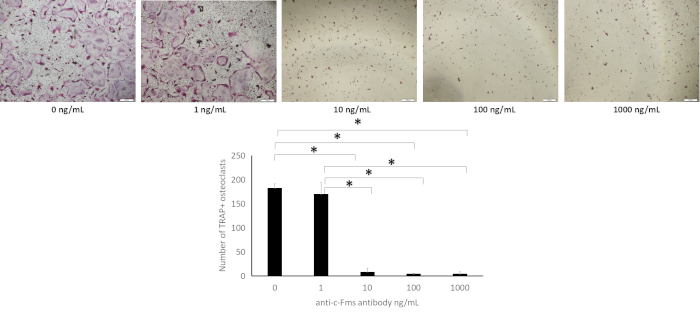
Figure 2: Effect of anti-c-fms antibody on TNF-α-induced osteoclast formation. Microscopic observation of osteoclast precursors that were treated with M-CSF, TNF-α and various dose of anti-c-fms antibody (0, 1, 10, 100 and 1,000 ng/mL) for 4 days. Cells were fixed and stained with TRAP staining. Number of TRAP-positive cells cultured with M-CSF, TNF-α and various concentrations of anti-c-fms antibody (0, 1, 10, 100 and 1,000 ng/mL) for 4 days. Cells were fixed and stained with TRAP staining. The number of TRAP-positive cells was assessed. Results were expressed as meanv ± S.D. of four cultures. Statistical differences were detected by using Scheffe's tests (n = 4; *p < 0.01). Scale bars = 200 µm. Please click here to view a larger version of this figure.
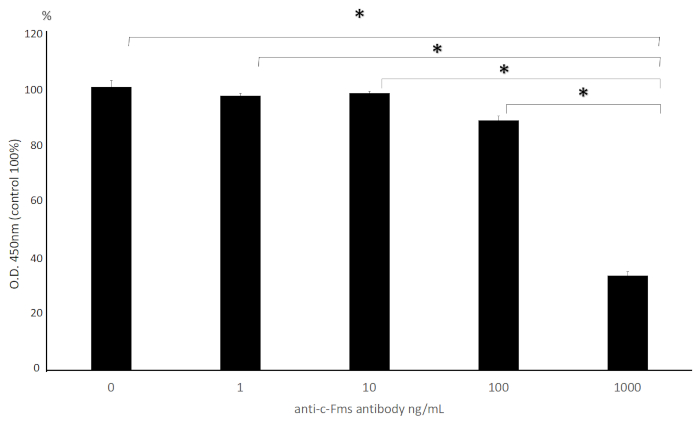
Figure 3: Effect of anti-c-fms antibody on proliferation of osteoclast precursor. Cell viability of osteoclast precursors that were treated with M-CSF and various dose of anti-c-fms antibody (0, 1, 10, 100 and 1,000 ng/mL) for 3 days. Cell counting kit-8 was used to measure viability. Data are presented as a percentage to compare the relative activity of the wells containing M-CSF+anti-c-fms versus the wells containing M-CSF alone and expressed as mean ± S.D. of four cultures. Statistical differences were detected by using Scheffe's tests (n = 4; *p < 0.01). Please click here to view a larger version of this figure.
