TERPS is used to distinguish between mono- and polysynaptic contributions in synaptic connections between neurons. While weak stimulation of a cell may be used to test direct connections, driving greater presynaptic activity often recruits polysynaptic connections (Figure 1A). TERPS works by co-expressing the TTX-insensitive sodium channel NaChBac and an optogenetic activator, and testing connections in the presence of TTX to eliminate polysynaptic connections (Figure 1B). TTX effectively blocks action potentials in the Drosophila brain, making it a suitable preparation for TERPS (Figure 2A). Transgenic expression of the NaChBac sodium channel rescues excitability in neurons and results in a large plateau potential (Figure 2B).
Local interneurons (LN) in the antennal lobe show robust responses to olfactory stimulation (Figure 3A). However, because individual EPSPs are not easily resolved at the LN soma, it remains unclear if such responses arise directly from olfactory receptor neuron input (ORN) or indirectly input via projection neurons (PN) (Figure 3B). By using TERPS, ORNs shown here, do indeed make direct synaptic connections with LNs (Figure 3C).
A unique feature of TERPS is its ability to specifically resolve extra-synaptic volume transmission that may occur with either GABA or neuromodulators such as serotonin. Stimulation of a serotonergic neuron (the CSDn) in the Drosophila antennal lobe results in a mixture of excitation and inhibition in a local interneuron (Figure 4A and Figure 4B). The excitation is meditated by the excitatory transmitter acetylcholine and the inhibition is caused by serotonin (Figure 4C and Figure 4D). TERPS can be used to determine if the connections are monosynaptic by eliminating polysynaptic connections (Figure 4E). In TTX, a strong inhibition is still observed in the LN, suggesting it arrives from directly from the activation of a specific serotonergic neuron (Figure 4F and Figure 4G). This connection is blocked by the serotonin antagonist methysergide. The excitatory synapse mediated by acetylcholine was eliminated in TTX, suggesting it arose from polysynaptic sources.
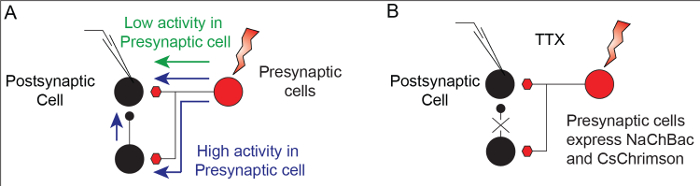
Figure 1: TERPS eliminates polysynaptic connections and isolates monosynaptic inputs. (A). A schematic diagram showing that increasing presynaptic activity can recruit polysynaptic connections. (B). A diagram illustrating how TERPS can remove polysynaptic contributions by using TTX to eliminate spiking activity in neurons. Excitability is exclusively restored in one population of neurons that can then be excited optogenetically. Please click here to view a larger version of this figure.
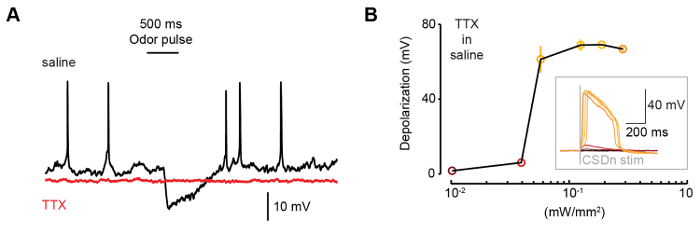
Figure 2: NaChBac restores neuronal excitability in Drosophila neurons. (A) The application of 1 µm TTX abolishes spiking in Drosophila neurons. Both spontaneous activity and odor-evoked inhibition are eliminated in TTX. (B) NaChBac and csChrimson are expressed in the CSDn and the brain is exposed to TTX. csChrimson stimulation depolarizes the CSDn and after a threshold is crossed, there is large non-linear increased in the CSDn membrane voltage. This rapid depolarization constitutes a plateau-like potential (inset). Each color across simulation intensities corresponds to the same color in the voltage inset. This figure has been modified from Zhang and Gaudry 20169. Please click here to view a larger version of this figure.
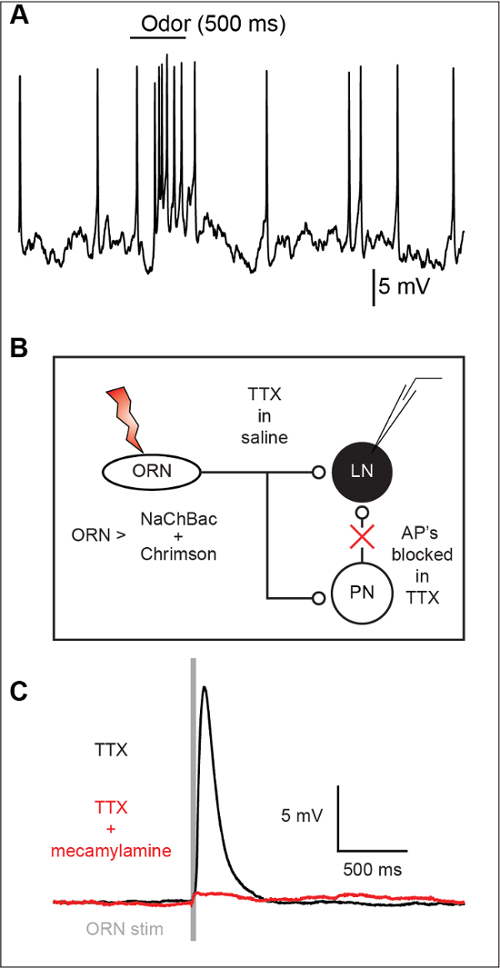
Figure 3: TERPS reveals monosynaptic connections between ORNs and LNs. (A) A sample recording showing an odor response in an LN. This response may be mono- or polysynaptic in nature. (B) TERPS can be tested at the ORN to LN synapse. TTX is used to block action potentials and excitability in all cells in the preparation. Excitability is exclusively restored in the ORNs via the NaChBac sodium channel. The ORNs can then be excited with channelrhodopsin to elicit synaptic release. Synaptic events are measured post-synaptically using whole-cell recordings. (C) TERPS reveals that the ORN to LN synapse is monosynaptic. TERPS can also be combined with pharmacology to show that the synapse is cholinergic and blocked by the nicotinic receptor antagonist mecamylamine (200 µM). The vertical gray bar indicates the timing of ORN stimulation. This figure has been modified from Zhang and Gaudry in 20169. Please click here to view a larger version of this figure.
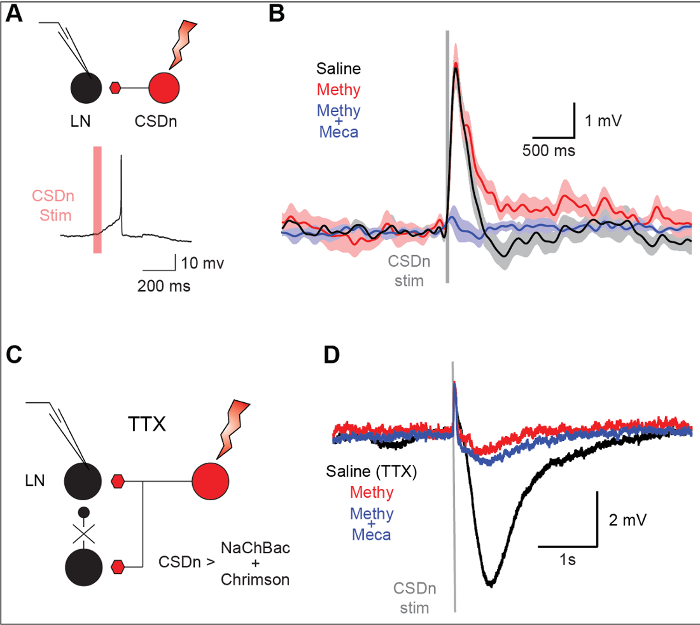
Figure 4: TERPS is sensitive to bulk or volume transmission release from modulatory neurons. (A) The stimulation of the CSDn results in an action potential from a recorded LN in the Drosophila antennal lobe. (B) The LN is hyperpolarized so that CSDn stimulation results in a subthreshold activity. The LN response consists of a fast depolarization followed by a slower hyperpolarization. The gray bar denotes the time of CSDn stimulation. Methysergide (50 µM), a broad 5-HT receptor antagonist, blocks the slow hyperpolarization but has no effect on the depolarization. Mecamylamine blocks the fast depolarization suggesting that it is cholinergic in nature. (C) TERPS can be used to resolve which chemical components of the CSDn to LN synapse are mono- versus polysynaptic. (D) The CSDn is stimulated in the presence of TTX and postsynaptic LN responses are measured in whole-cell recordings. The hyperpolarization persists in TTX suggesting it is monosynaptic in nature. This hyperpolarization is also blocked by methysergide, consistent with serotonergic transmission. The brief depolarization that remains in TTX is likely mediated by electrical gap coupling, as it not blocked by nicotinic antagonists. This figure has been modified from Zhang and Gaudry in 20169. Please click here to view a larger version of this figure.
Supplementary File 1: Drosophila Foil. Please click here to download this file.
Supplementary File 2: Physiology Chamber. Please click here to download this file.
