With respect to experimental technique, the most critical component of the protocol is keeping all the samples cold. Trypanosomes very rapidly internalize antibody bound to their surface9, but this process is motility dependent, and does not influence the assay as long as the cells are kept at 4 °C. All samples should always be kept on ice, and pipetting should be done quickly to minimize exposure to the 25 °C lab environment. Cold HMI-9 with serum should be available at the start of the experiment and should be kept on ice throughout. Isolation of the trypanosomes by running them over the column must be done in a cold room, as it takes 5 – 7 min to run a 1 ml sample through the column. If there are more samples than magnetic separation units available, magnetic isolation can be done in sequence, as long as samples not being actively separated are kept on ice.
It is also critical that samples be very well mixed after the last wash following incubation with the microbeads and prior to isolation on the magnet. This can be accomplished by aggressive flicking of the centrifuge tube or light vortexing. No pellet or clumps of cells should be visible prior to addition of the sample to the separation column, which can be checked by looking at the light microscopy level. If clumps are present, the level of contamination in the flow-through sample of cells expressing the dominant VSG is usually much higher than desirable. When performing the calculation for the number of switchers in the sample at the end of the protocol, it is important to take into account whether all the flow-through was used for flow cytometric analysis, or whether half was used. If only half was used, the total number of switchers calculated should be multiplied by 2.
When removing the supernatant during the wash steps, it is not necessary to pipette off every last drop, and typically 15 – 25 µl are left surrounding the pellet of cells. This is especially critical during the final stages of staining and washing, because at this point pellets are rarely seen in the flow-through samples because there are so few switched cells.
While the final anti-VSG stain is typically performed with purified, fluorophore-labeled anti-VSG antibody, the first anti-VSG stain prior to magnetic isolation can be done with purified antibody or serum. We have also used bioreactor supernatant derived from hybridomas expressing anti-VSG antibodies. Regardless, the material used for the initial stain must be titrated to ensure good separation of cells that do or do not express the dominant, starting VSG.
It is typical for the population of cells in the flow-through to be contaminated with cells expressing the dominant VSG. While this contamination should be minimal, in our experience it is rare for the population to be completely devoid of cells expressing the dominant VSG. It is also important to note that cells that have gone through the isolation procedure do not stain as brightly with anti-VSG antibody as those that have not been through the procedure. We hypothesize that two things could account for this difference. The first is that spinning trypanosomes leads to shedding of VSG, and there are many spins involved in the isolation procedure. This would be expected to decrease the intensity of the signal for the final anti-VSG stain. The second is that the first step of the protocol involves staining with a primary anti-VSG antibody. If the epitopes recognized by the antibody used in this first step are the same as or occlude the epitopes recognized by the antibody in the final stain, one would expect the intensity of the signal to be lower than if only one antibody were used. For this reason it is important to titrate the antibody that is used in the final step, such that the signal from a positively stained cell is roughly two orders of magnitude higher than for a negatively stained cell. This way, even if the intensity of signal from a positively staining cell is decreased in cells that have undergone the isolation procedure, these positive cells will still be well separated from the negative cells on the flow cytometry plot. If auto-compensation is being used, single stain control samples should be prepared. Antibodies that we have generated are available for purchase from the Memorial Sloan Cancer Center monocolonal antibody facility. We have had the best success with using unlabeled monoclonal IgG antibody. We have carried out the protocol with polyclonal VSG antibodies, but polyclonal antibodies can occasionally recognize other VSGs so monoclonal antibodies are preferred. VSG identity can be determined by sequencing VSG cDNA generated from amplification of conserved sequences in the VSG 3'UTR and the spliced leader. A particular antibody can be tested for specificity on previously isolated switched cells who's VSG identity has been determined as just described.
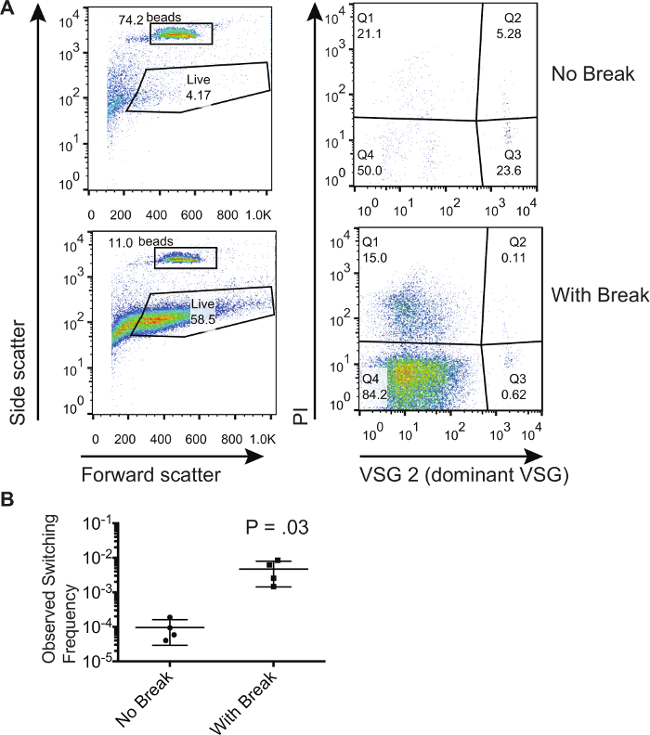
The protocol presented here has a number of advantages. It can be used both to isolate switchers for later analysis and to quantify the number of switchers in a given population. It also can be used to isolate a particular variant of interest, as long as an antibody is available for that variant. It is also possible to carry out this protocol using trypanosomes isolated from animals, provided that red blood cells are eliminated using anti-Ter119 coated-magnetic beads according to the manufacturer's protocol. We have not tested the lower limit for the number of trypanosomes required, but we have successfully carried out the protocol using 2.5 – 10 million trypanosomes from 250 µl of blood. Finally, the protocol can be used in conjunction with fluctuation analysis to obtain the frequency of switching in a given population.
The isolation procedure is quite fast, taking 3 – 4 hr from start to finish depending on the number of samples, and thus is more efficient than previous methods that required prior immunization of mice or specialized strains containing drug resistance markers. The method is limited by the fact that antibodies against the starting VSG are required, however. Without such reagents, an alternate method such as VSG-seq might be a more appropriate choice to gauge which VSGs are being expressed in a given population10.