All animal experiments were carried out in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals (NIH Publications No. 8023, revised 1978) and with the Animal Care and Use Committee of the Institutional Animal Care and Use Committee of KIST (Seoul, Korea).
1. Animal Preparation
- Prepare a group of 7-week-old male ICR mice (or C57BL/6).
- Let the mice go through an acclimation process for 3-7 days after transport to restore homeostasis. In general, place a group of 4-5 mice in each cage and maintain them at 22-23 °C and 40% humidity, with a 12/12 hr light/dark cycle. Provide water and food ad libitum.
Note: The acclimation process is critical to allow recovery from the stress of shipping and adaptation to new housing, food, water, and smell as well as a new cage and light cycle. In this experiment, mice were housed in Individually Ventilated Cages (IVCs) located in the Specific Pathogen Free (SPF)-grade facility - Weigh each mouse and exclude subjects whose weight deviates ± 10% from the average.
- Divide the subjects into two groups: a vehicle-injected group (Aβ(-)) and an Aβ-injected group (Aβ(+)). If the experiment is to test the efficacy of a particular drug candidate, divide the mice into four groups: (1) Aβ(-)/drug(-), (2) Aβ(-)/drug(+), (3) Aβ(+)/drug(-), and (4) Aβ(+)/drug(+).
2. Aβ Peptide Preparation
- Treat the Aβ peptides, derived through chemical synthesis5 or from commercial sources, with pre-chilled 1,1,1,3,3,3-hexafluoro-2-propanol (HFIP) to dissolve any aggregates and to make the peptides homogenous6.
- Sonicate the Aβ/HFIP solution in a water-bath sonicator for 5 min (at RT), gently vortex, and incubate the solution for 30 min at RT. Remove the HFIP completely via evaporation with nitrogen and store the homogenous Aβ at -80 °C until use.
- Prepare the Aβ monomers: make 1 mM Aβ dimethylsulfoxide (DMSO) stock and dilute it 10-fold in phosphate buffered saline (PBS) (100 µM Aβ, 10% DMSO, 90% PBS.)
- Prepare Aβ oligomers.
- Incubate the Aβ monomer solution (containing Aβ(1-42) or Aβ(1-40)) (step 2.2.) at 37 °C for 3 or 7 days, respectively.
- Place the Aβ solution in a sealed tube in a zippered bag containing a wet paper towel to provide a humidified environment to prevent water evaporation during the incubation period.
- Confirm prepared Aβ oligomers via sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE) with photo-induced cross-linking of unmodified proteins6.
- Prepare the control PBS solution containing 10% DMSO for the vehicle ICV injection.
3. Syringe Preparation
- Prepare a microsyringe with a 26 G stainless steel needle for ICV injection.
- Sterilize the syringe in autoclave with all the other apparatus that will be used for surgical procedure. After the autoclave, place the apparatus under the UV light for 20 min.
- Wrap the needle of the microsyringe with Parafilm to adjust its length to 3.8 mm to enable maximum accuracy in the depth of the injection into the brain (Figure 1A). (For B6 mice, adjust syringe needle to 3.6 mm).
Note: Compression of the Parafilm will lead to the depth of the injection exceeding 3.6 mm ventral. To prevent the Parafilm from being compressed during needle insertion, densely wrap the needle multiple times. - Wash the inner space of the syringe with 70% ethanol, twice.
- Sonicate the syringe and the needle in a water-bath sonicator for 30 min at RT.
- Wash the inner space of the syringe with 70% ethanol, twice.
- Rinse out the syringe with distilled water and dry it completely in a fume hood for longer than 30 min. Leave the clean syringe under a UV light for 20 min before starting the Aβ injection.
4. Aβ-injected Mouse Model Preparation
Note: Clean the operating field with a disinfectant to maintain sterile conditions and sterilize all surgical instruments for ICV injection using 70% ethanol and UV exposure.
- Anesthetize the mouse by intraperitoneal injection of a mixture of xylazine (20 mg/kg) and tiletamine·HCl/zolazepam·HCl (80 mg/kg) before the ICV injection. Place the mouse on a warm mat to maintain its body temperature while anesthetized and confirm adequate anesthesia by assessing the foot-pinch response.
- Apply sterile PBS drops to both eyes to prevent corneal dryness during the procedure (Figure 1B).
- Prepare two mirrors by drawing multiple straight vertical lines on their surfaces in case of difficulty injecting the syringe perpendicularly.
- Place one mirror (M1) right next to the body of the mouse and the other (M2) in front of the head. Keep M1 parallel to the imaginary midline between the two eyes of the mouse and arrange M2 perpendicular to M1, so that the two planes form a 90° angle (Figure 1C). Place M1 on the left side of the mouse if the researcher is right-handed. Place M1 on the right side of the mouse if the researcher is left-handed.
- Spray 70% ethanol onto the middle of the forehead of the mouse and rub it with dry cotton swabs. Do not wet too large of an area of the forehead to avoid decreasing the body temperature due to evaporation of the alcohol.
- Wipe the same area on the forehead with 2% chlorhexidine solution using a clean cotton swabs. Repeat scrubbings with alcohol and chlorohexidine for total of three times each.
Note: If necessary, shave hair on the forehead of the mouse in order to minimize the possibility of contamination - Locate bregma.
- Using the thumb and index finger, tightly hold down the skin of the forehead, directly above the two eyes to the degree that both eyes protrude. Drag the skin back to make the forehead taut and minimize skull movements under skin.
- Form an invisible triangle by assigning three vertices: the two eyes of the mouse and the point where the thumb and index finger meet, the bregma. (Figures 1B, Red arrow: bregma)
- Locate the injection point with measuring tape: -1.0 ± 0.06 mm posterior to bregma, 1.8 ± 0.1 mm lateral to the sagittal suture, and 2.4 mm in depth (Figure 1D, blue stars: injection points in each hemisphere). (For B6 mice: -0.9 mm posterior, 1.7 mm lateral, and 2.2 mm in depth)
- Fill the syringe with 10 µl Aβ solution or vehicle with excess solution in order to avoid accidental air injection.
- Place the syringe at the injection point (described in protocol 4.7). Make the syringe perpendicular to the plane of injection point. Align reflected images of the syringe in both mirrors with the drawn lines on the mirrors from a fixed perspective (Figure 1C).
- Begin inserting the needle until the parafilm wrapping reaches the skin.
- Keep the hand holding syringe steady and use other hand to inject 5 µl of the Aβ solution or vehicle, slowly over 5 sec without pausing. Make sure the syringe remains perpendicular throughout. After completing the injection, wait 3 to 5 sec before removing the syringe for diffusion.
- Assuming the original triangle position, exert moderate pressure to protect the skull from any unnecessary movements. Remove the syringe without tilting.
- Place the Aβ-injected mouse on warm pad for recovery and maintain its sternal recumbency. Do not leave the mouse unattended until it regains consciousness and do not return the mouse to a cage containing non-surgical mice until it has fully recovered.
- Wash the syringe according to steps 3.3-3.6.
- Postoperatively monitor the mobility of the mice and check for any sign of infection or illness in the mice for 5-7 days.
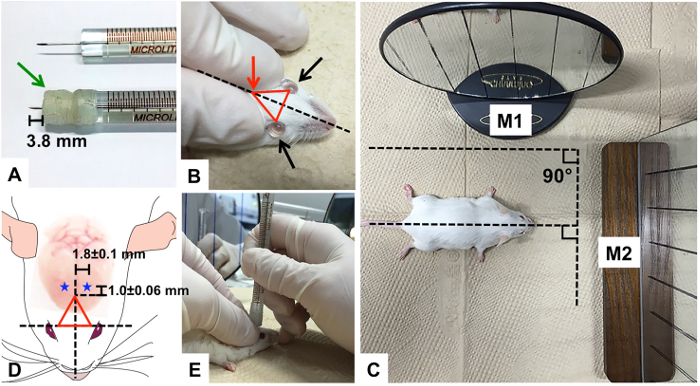
Figure 1. Aβ Injection into the ICV region. (A) Parafilm-wrapped syringe needle compared to an unmodified syringe needle; (B) PBS drops onto eyes to prevent dryness; the triangle formed on the forehead of mouse with thumb, index finger, and the two eyes of mouse as well as the imaginary midline equidistant from each eye; (C) two mirrors (M1 and M2) with multiple vertical lines drawn on the surfaces to assist perpendicular injection of the syringe needle; (D) the triangle with the injection points, indicated as blue stars (-1.0 ± 0.06 mm from bregma and 1.8 ± 0.1 mm sagittally) on each hemisphere of mouse; Green arrow: parafilm, Red arrow: bregma Please click here to view a larger version of this figure.
5. Confirmation of Aβ Injection by Y-maze and Brain Analyses
- Assess the working memory of Aβ-injected mice by Y-maze tests3,7,8
- Wait 3 days after the injection for the onset of memory deficits.
- Perform the Y-maze tests using a black plastic maze with three identical arms labeled A, B, and C, each of which branches out at a 120° angle from the center of the maze (width x length x height, 10 cm x 40 cm x 12 cm).
- Place a mouse at the start arm (arm A) and allow the subject to freely explore the maze for 8 min.
- Measure the number of entries into each arm and calculate the alternation rate of each subject3,9.
- Directly examine postmortem brains to check if the injection targeted the ICV region appropriately.
- Anesthetize the mouse by the same procedure as outlined in section 4.1. Then sacrifice the mouse by cervical dislocation.
- Decapitate the mouse and remove the skin to expose the skull using surgical scissors. Remove tissues and muscles from the cranium.
- Make cuts on the lambdoid suture (between the parietal and interparietal bones) without damaging the brain. Remove the occipital and interparietal bones, then the parietal and frontal bones. Remove the cranial bone using forceps and isolate the brain from the skull by lifting up the meninges.
- Wash the brain in chilled sterile saline and cut the ICV region of the brain in the coronal direction with a surgical knife (Figure 2).
- Confirm the injection region based on the trace of the needle insertion on the brain tissue (Figure 2). Check if the needle trace has reached one of ventricles. If the needle trace does not meet or pass through the ventricles, exclude that subject's data from analysis of the experimental results.
This section illustrates examples of the results that can be obtained by confirmation of Aβ aggregation and Y-maze assessment of memory deficits. Using the full-length Aβ(1-42) peptide of 42 amino acids, mixture of Aβ monomers, oligomers, and fibrils (Figure 3) was produced. Through the HFIP-induced monomerization step, relatively homogeneous monomers (Lane B) were obtained. After the 7 day incubation, diverse sizes of Aβ aggregates (Lane C) developed. Trimers and tetramers were the dominant species among the oligomeric forms of Aβ. Spatial working memory was assessed in the Aβ-injected mice via alternations in the Y-maze test. The sequence of arm choices and the number of total arm entries were recorded while each mouse was allowed to freely explore the maze. The more intact the cognitive ability, the more the mouse should have a tendency to enter the less recently visited arm, alternating its arm choice. The Aβ-injected mouse group showed significantly lower alternation rates, indicating the development of cognitive deficits (Figure 4).
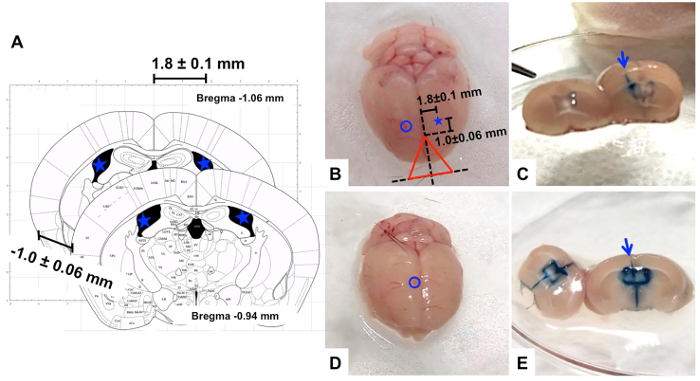
Figure 2. Examples of ICV injections (A) Illustration of brain sections in coronal direction representing the location of bregma10 and acceptable injection range (-1.0 ± 0.06 mm from bregma and 1.8 ± 0.1 mm sagittally). (B–C) a case of successful ICV injection (B: top view of the whole brain, C: coronal section showing lateral ventricles filled with blue dye); (D–E) a case of unsuccessful ICV injection (D: top view of the whole brain, E: coronal section showing the third and fourth ventricles filled with blue dye) Blue circle and arrow: injected site, Blue star: potential injection point. Please click here to view a larger version of this figure.
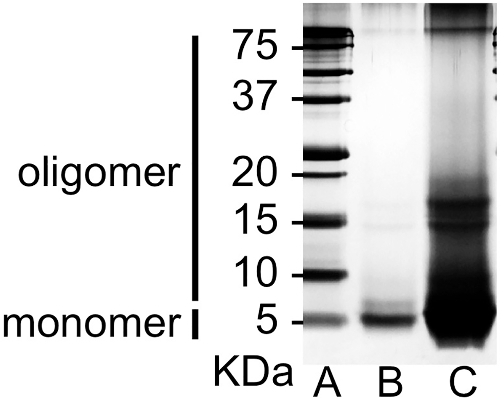
Figure 3. Confirmation of prepared Aβ species via SDS-PAGE. Aβ peptides were separated by SDS-PAGE with photo-induced cross-linking of unmodified proteins. Peptide bands were visualized by silver staining; lane (A) protein marker, lane (B) Aβ monomer (before the incubation), and lane (C) incubated Aβ species. Please click here to view a larger version of this figure.
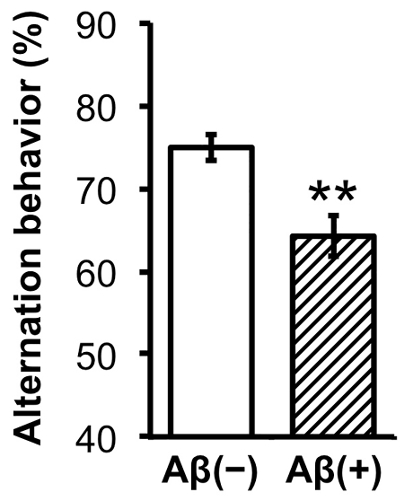
Figure 4. Y-maze behavioral tests. Percent alternation of the Aβ- or vehicle-injected mice groups on the Y-maze test. White solid graph: vehicle (Aβ(-), n = 10). Striped graph: Aβ-injected (Aβ(+), n = 9). Statistical analyses were performed with one-way ANOVA followed by Bonferroni's post-hoc comparisons (*P <0.05, **P <0.01, ***P <0.001). The error bars represent the SEMs. Please click here to view a larger version of this figure.
