Nuclear staining of thick specimens. The protocol herein described allows for the achievement of homogeneous staining of deep structures in whole embryos. Developing lens nuclei form 48 hpf zebrafish embryos are homogeneously labeled (Figure 1).
Methyl green DNA staining allows discrimination of cell cycle dependent morphological differences such as the identification of mitotic figures or apoptotic nuclei (Figure 2A). Subnuclear structures at high resolution are also evident as shown for epidermal nuclei of zebrafish embryos (Figure 2B).
Methyl green is resistant to photobleaching under high intensity irradiation for prolonged periods of time. When irradiated with 633 nm laser light at the same intensity, methyl green nuclear staining does not bleach out as it is the case of another DNA-specific stain sharing similar spectral characteristics, TO-PRO-34. A confocal plane of the retina of a whole-mounted zebrafish embryo was selected and zoomed in with a 63x 1.4NA objective to achieve high irradiation. When zoomed out, the specimens stained with TO-PRO-3 showed bleaching within 60 seconds of continuous irradiation (Figure 3, upper panels), while the specimens stained with methyl green showed no visible evidence of photobleaching even at 120 seconds in the same conditions (Figure 3, lower panels). Images acquired in LASAF software were further processed (brightness/contrast; 3D reconstruction; maximum intensity projection) using Fiji open source software (http://fiji.sc/).
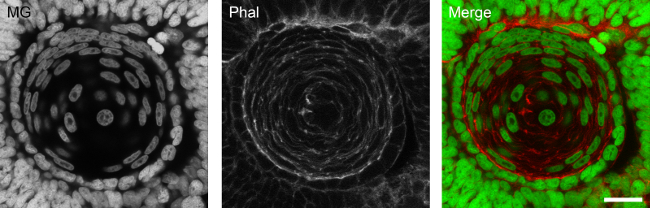
Figure 1: Homogeneous nuclear staining of deep structures with methyl green. Developing lens of a 48 hpf zebrafish embryo stained with methyl green (MG) in a z-projection of 20 confocal planes. Embryos were counterstained for F-actin with TRITC-conjugated phalloidin (Phal). Scale bar: 15 µm.
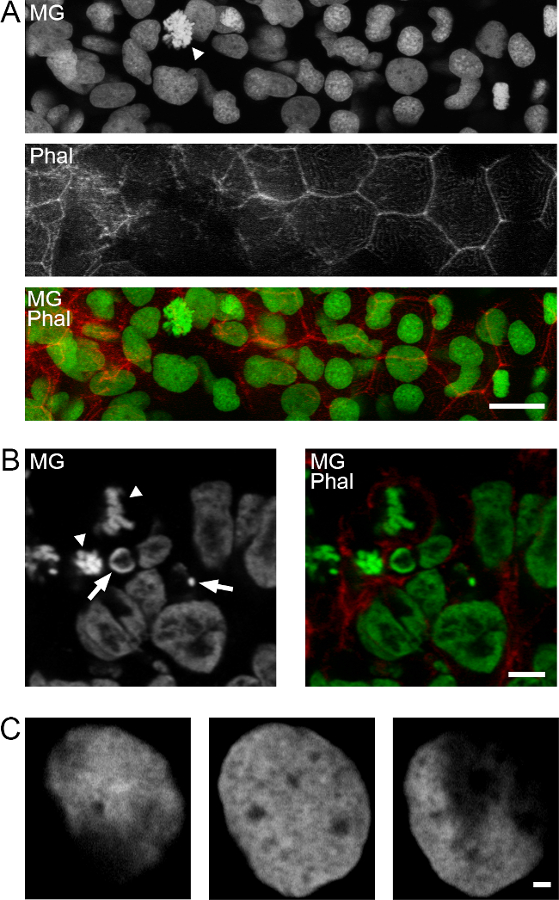
Figure 2: Nuclear morphology and subnuclear resolution with methyl green staining. A) Different nuclear morphologies in the epidermis of a 48 hpf zebrafish embryo. MG, methyl green. Specimens were counterstained for F-actin with TRITC-phalloidin. Arrowhead: mitotic cell. Maximum-intensity projection of 10 confocal planes. B) Chick embryo neural plate labeled in the same way, displaying mitotic cells (arrowheads) and pyknotic nuclei (arrows). Single confocal plane. C) Three single confocal planes of a zebrafish embryo epidermal nucleus at high magnification, showing a high degree of subnuclear resolution. Scale bars: A, 15 µm; B, 5 µm; C, 1 µm.
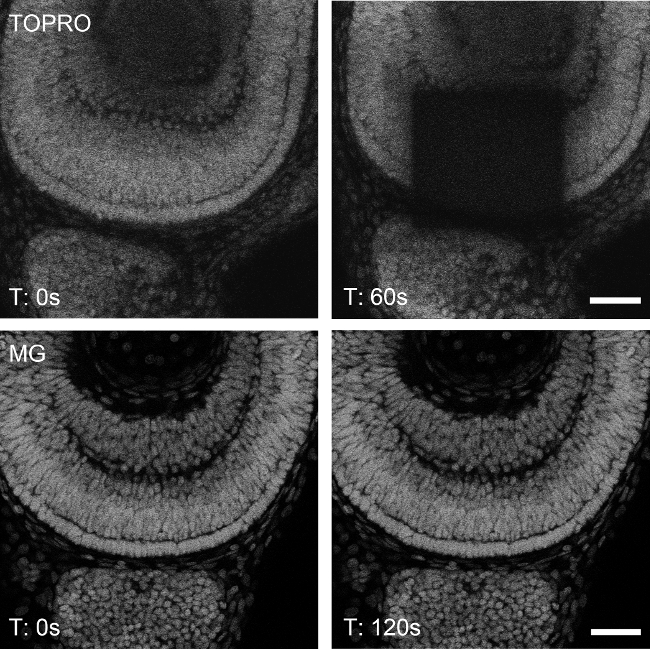
Figure 3: Photostability of methyl green under continuous excitation. TO-PRO-3-stained nuclei of a 48 hpf zebrafish retina was bleached upon 60 sec of continuous excitation at 633 nm (upper panels). Under the same conditions, a methyl green-stained embryo (MG) did not show perceptible bleaching, even when duplicating the exposure time (lower panels). Bleaching and acquisition was made by scanning in single planes, at 8,000 Hz, 30% laser power and line (x2) and frame (x4) averaging. Scale bar: 30 µm.
| Wavelength (nm) | Percent emission |
| 640 | 21.37 |
| 643 | 27.5 |
| 646 | 36.69 |
| 649 | 44.47 |
| 652 | 53.92 |
| 655 | 61.82 |
| 658 | 67.86 |
| 661 | 83.2 |
| 664 | 85.22 |
| 667 | 89.45 |
| 670 | 87.33 |
| 673 | 92.3 |
| 676 | 100 |
| 679 | 97.41 |
| 682 | 90.34 |
| 685 | 83.32 |
| 688 | 78.37 |
| 691 | 76.09 |
| 694 | 73.95 |
| 697 | 67.36 |
| 700 | 64.4 |
| 703 | 61.26 |
| 706 | 56.39 |
| 709 | 55.06 |
| 712 | 49.56 |
| 715 | 46.12 |
| 718 | 41.4 |
| 721 | 38.8 |
| 724 | 35.97 |
| 727 | 33.27 |
| 730 | 30.95 |
| 733 | 28.78 |
| 736 | 26.28 |
| 739 | 24.96 |
| 742 | 23.3 |
| 745 | 21.12 |
| 748 | 19.86 |
| 751 | 17.47 |
| 754 | 16.14 |
| 757 | 14.21 |
| 760 | 12.45 |
| 763 | 11.49 |
| 766 | 9.97 |
| 769 | 7.55 |
| 772 | 7.75 |
| 775 | 6.46 |
| 778 | 5.76 |
| 781 | 4.38 |
| 784 | 2.6 |
| 787 | 2.75 |
| 790 | 2.27 |
| 793 | 1.52 |
| 796 | 0 |
Supplementary material 1: Emission spectrum of methyl green nuclear staining under 633 nm excitation.
