Exemplary isolation of single cells was performed from two mandibular incisors from one 6-week-old C57BL/6 mouse male. Following this protocol, a single-cell suspension was prepared, and subsequently, single-cell sequencing was performed. The prepared single-cell suspension was analyzed and sorted using FACS (Figure 2). Firstly, the FSC-A (forward scatter, area) and SSC-A (side scatter, area) plotting was applied, and an appropriate gating strategy was used to select a population with expected size and granularity to filter our cell debris and cell doublets or aggregates (Figure 2A). This selected population (P1), counting 38% of all events, was further used, and FSC-A and FSC-H (forward scatter, height) parameters were applied to remove the remaining cell doublets (Figure 2B). The population without cell doublets (P2) counting 95% of P1 can be subsequently used for scRNA-seq. Alternatively, additional gating can be used to select the population of interest (e.g., expression of fluorescent proteins or live/dead staining). To check the number of live/dead cells in the final suspension, the PI (propidium iodide) staining was performed (Figure 2C). The P3 population containing PI– (living) cells was 98.4% out of the parent P2 population and 35.5% out of the total events. The total number of filtered out, dead (PI+) cells was 1887.
The final number of cells obtained without viability staining suitable for RNA-seq (P2) counted 118,199 cells from two mouse incisor pulps. This means the number of almost 60,000 living cells from one mandibular incisor.
To clarify the number of immune cells in the final single-cell suspension, two approaches were used. Firstly, the CD45 antibody staining and subsequent FACS analysis were used. As a complementary method, the total number of immune cells (CD45+) in scRNA-seq data was analysed. FACS analysis showed 14.44% of CD45+ cells (13.20% alive and 1.24% dead) (Figure 3A). Analysis of scRNA-seq data showed 10.90% of CD45-expressing cells (Figure 3B). The decrease of CD45+ cells in scRNA-seq data can be caused by additional thresholding during scRNA-seq analysis.
These representative data on the example of mouse incisor show that the given protocol in combination with strict gating strategy is efficient in obtaining a high number of cells out of a single mouse tooth without the necessity of additional use of viability staining. The ratio of immune (CD45+) cells was minor (13.2%). Moreover, it was previously shown that the immune cells are essential in maintaining tooth homeostasis, so removing them from scRNA-seq analysis during the FACS would be counterproductive in some applications.
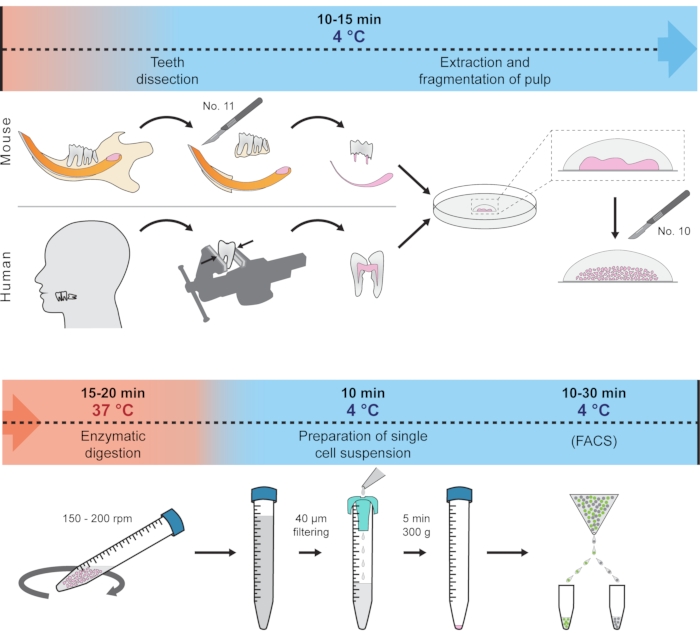
Figure 1: Schematic representation of the protocol. Different steps, including temperature conditions and expected time, are represented to prepare single-cell suspension from mouse and human teeth. Please click here to view a larger version of this figure.
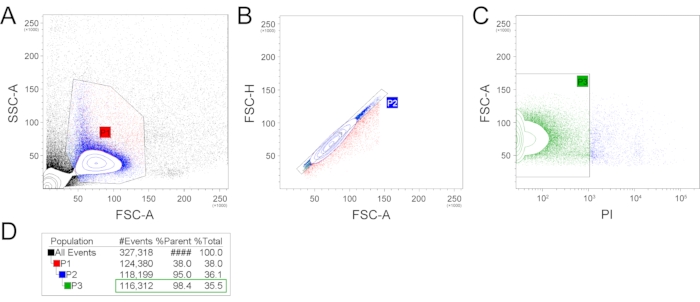
Figure 2: Example of the gating strategy. FSC-A and SSC-A gating was used to produce the P1 gate, reflecting the cell population with expected size and granularity and filtering out the cell and extracellular matrix debris and most large events (A). Subsequently, the P1 population was plotted in FSC-H and FCS-A plot, which filtered out cell doublets (B). This P2 population was then analyzed for the presence of dead cells by propidium iodide (C). The number of events/cells per gate are represented in (D). (FSC-A – forward scatter, area; SSC-A – side scatter, area; FSC-H – forward scatter, height; PI – propidium iodide). Please click here to view a larger version of this figure.

Figure 3: Quantification of the immune cells. Quantification of the immune cells was performed by FACS analysis of cells stained with anti-CD45 antibody and Live/Dead analysis using propidium iodide staining (A). Further quantification of immune cells was performed during scRNA-seq analysis (B). (CD45-APC – anti-CD45 allophycocyanin conjugated antibody; PI – propidium iodide; t-SNE – t-distributed stochastic neighbor embedding). Please click here to view a larger version of this figure.
Supplementary Figure 1: Overview of the mandible dissection process. Dashed lines illustrate suggested cuts. TMJ – temporomandibular joint, m. masseter – musculus masseter. Please click here to download this File.
Supplementary Table 1: The compositions of the solutions used in the study. Please click here to download this Table.
