Results of microsatellite analysis using F0 and F1 generations showed that inbreeding crosses were produced successfully (Figure 4)6. As a result of inbreeding crosses, mated queens were obtained within one month of establishing the experimental crossing colonies. A quarter (27.1 ± 8.91% SD) of all offspring (F2) from the inbreeding crosses was male, while the remainder was female (workers and a queen)6. QTL mapping using the offspring from inbreeding crosses showed that males produced by inbreeding crosses were diploid and homozygous at two CSD loci (CSD1 and CSD2 in Figure 5), while females (workers) produced from inbreeding crosses were diploid and heterozygous at least at one CSD locus6.
Dissection of haploid males revealed testes and sperm, as expected (Figure 6A–6C). However, in diploid males, sperms were never observed, suggesting that males produced in inbreeding crosses are sterile in V. emeryi6. In addition, testes of diploid males failed to develop (Figure 6D–6E).
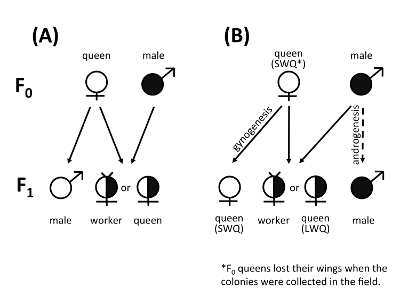
Figure 1. Typical reproductive system in (A) Hymenoptera and the atypical reproductive system involving androgenesis and gynogenesis in (B) V. emeryi. Typically, females (workers and queens) develop from fertilized diploid eggs, and males develop from unfertilized haploid eggs containing half of the maternal genome (A). In V. emeryi, sterile workers and a few long-winged queens (LWQ) develop from fertilized diploid eggs, while short-winged queens (SWQ) develop with nearly complete maternal genomes from unfertilized diploid eggs (gynogenesis). Males never inherit maternal genomes but are clones of their fathers (androgenesis) (B). This figure has been modified from [Miyakawa et al. 2018]7. Please click here to view a larger version of this figure.

Figure 2. Experimental set up of V. emeryi colonies. After field collection, colonies are transferred into an artificial plaster nest and kept in the laboratory. A large plaster nest (left) is prepared for maintain collected colonies, whereas a smaller plaster nest (right) is prepared for experimental inbreeding crosses. Please click here to view a larger version of this figure.

Figure 3. New V. emeryi reproductives emerge during the reproductive season. Mature and well-fedcolonies tend to produce long-winged queens (LWQ) with the parental genome in addition to short-winged queens (SWQ) which bear only the maternal genome (Figure 1B). Photo courtesy of Mr. Taku Shimada. Please click here to view a larger version of this figure.
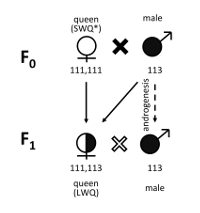
Figure 4. Design of inbreeding crosses and microsatellite genotypes of F0 and F1 generations. Using 11 microsatellite markers developed in previous studies6,8,9, females and males of the parental generation (F0) showed different genotypes. The genotypes of females and males used for experimental crosses (F1) inherited the parental and paternal genotypes, respectively, indicating that females were crossed successfully with their brothers, with which the females shared half their genomes. Numbers indicate lengths of PCR products at microsatellite locus L-5, which is one of the markers used for genotyping6,8,9,10. This figure has been illustrated according to the data from [Miyakawa and Mikheyev 2015]6. Please click here to view a larger version of this figure.
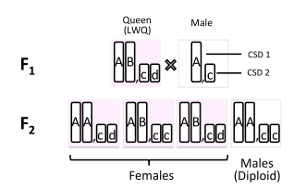
Figure 5. Allele patterns of two CSD loci (CSD1 and CSD2) in offspring produced by inbreeding crosses. Proportion of diploid males (about 25%) and QTL mapping using offspring produced by sib-mated queens suggest the existence of two CSD loci in V. emeryi. Females are heterozygous in at least one of the two CSD loci whereas males are homozygous at all loci. Genotypes are represented by letters of the alphabet. This figure has been modified from [Miyakawa et al. 2018]7. Please click here to view a larger version of this figure.
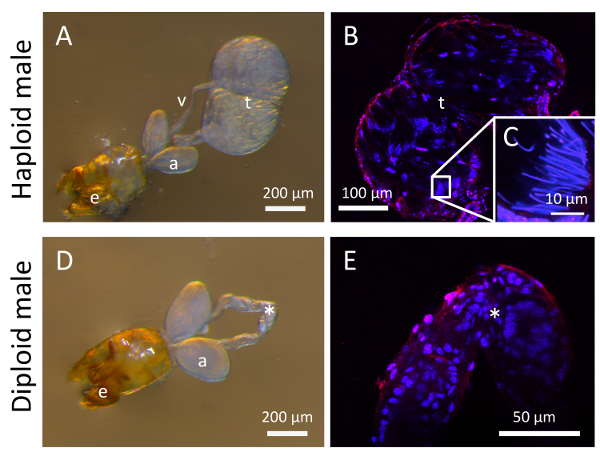
Figure 6. Male internal reproductive organs of androgenetic haploid and diploid males in V. emeryi. Morphologies of testes and other internal reproductive organs dissected out from the androgenetic haploid males (A). Sperm (fibrous tissue) could be seen in testes of haploid males (B and C). Blue color marks nuclei stained by DAPI, and red color marks F-actin stained by Tetramethylrhodamine (TRITC)-conjugated phalloidin in B, C, and E. (a) accessory glands; (t) testes; (v) vas deferens; (g) external genitalia. Morphologies of internal reproductive organs dissected out from the diploid males (D). Testes and sperm of diploid males were never observed (D and E, N >> 30). Please click here to view a larger version of this figure.
