Paraquat has been shown in numerous animal models, such as mice19, rats20 and fruit flies22, to induce neurological degeneration consistent with Parkinsonian-like pathology. We have previously reported that the anti-inflammatory antibiotic minocycline, when cofed with paraquat, results in extended survival, reduced production of reactive oxygen species and rescue of dopaminergic neuron death25. Therefore, as minocycline acts to suppress inflammation, we began investigating NOS, a key protein in the activation of the inflammatory response, and its role in modulating paraquat toxicity.
A paraquat toxicity curve was performed to establish an effective lethal dose and toxic treatment range, using paraquat concentrations between 1.25 and 10 mM (Figure 2A). We also assayed the effects of paraquat concentrations of 20 mM and 40 mM, but found that toxicity was too acute at these higher concentrations to accurately assess cellular responses to this oxidative stressor, and therefore, we have not included the results for these concentrations in this report. Using L-NAME, a competitive NOS inhibitor, and D-NAME, the inactive isomer of NAME, we cotreated adult male flies with paraquat and one of the NAME isomers. Cotreatment of flies with paraquat and L-NAME resulted in a significant rescue of lifespan truncation caused by paraquat ingestion, while flies cotreated with paraquat and D-NAME showed no improvement in survival (Figure 2B), supporting the hypothesis that suppressing inflammation through the inhibition of NOS enhanced survival of paraquat-treated flies.
As further validation of a NOS-mediated paraquat response, we detected changes in NOS protein levels directly after paraquat exposure. NOS protein levels increased in a paraquat concentration-dependent manner (Figure 3A). When treating flies with 10 mM paraquat over exposure durations ranging from 6-30 hr, we observed an initial increase, then a decrease as exposure time increased (Figure 3B), consistent with the patterns of nitrite levels observed in our variant of the Griess assay (Figure 5B). These results are consistent with our reports that paraquat treatment causes induction of NOS and that inhibition of NOS or treatment with L-NAME or minocycline provides partial protection17,25.
Figure 4A demonstrates a linear relationship between the increase in paraquat concentration and the magnitude of the inflammatory response as defined by the secretion and detection of NO. Under basal levels, heterozygous Punch mutants secreted slightly lower levels of NO, a variation too indiscernible to be determined using previous detection methods. When fed paraquat, a pronounced increase in NO levels were observed in the Punch mutant, however, significantly less than the wildtype treated flies (Figure 4B). The relative values of the wildtype and Punch flies can be accurately and reproducibly quantified, though the variation at untreated levels were extraordinarily subtle.
When working with toxins or chemicals, it is vital to establish both a time-of-exposure curve as well as a concentration-based toxicity curve, since these conditions can dramatically alter the sensitivity of the assay. For example, treatment with 5 mM paraquat results in maximum NO detection after 24 hr of exposure (Figure 5A), while exposure to 10 mM paraquat resulted in more rapid induction (maximum levels at 12 hr), but also more rapid decay of activity during extended exposures (Figure 5B). Although a higher concentration with quick induction might seem preferable, a maximized response accelerates death of both neurons and possibly hemocytes to the point that reproducibility may become difficult.
NO molecules are unstable and highly reactive. A successful assay must therefore include incubation conditions that provide an optimal balance between continued induction of NOS and rapid turnover of NO. In order to define optimal incubation conditions, we assessed the effect of the time that dissected brains were incubated in the culture medium prior to adding the Griess reagent. We found that a 6 hr incubation at room temperature produced maximum nitrite levels in the subsequent Griess reaction (Figure 5C) .We expect that variations in the incubation period may be needed to optimize detection for various models of inflammation. Careful establishment of all optimal parameters for the particular organism, genotype, and tissue being assayed, will be essential to achieve reliable and accurate results since the process is highly dynamic. In particular, genetic variants and transgene expression models are expected to require substantial optimization, particularly with respect to developmental stage or age of adults.

Figure 1. Drosophila adult brain. Light microscopy of dissected adult male brain 2-3 days posteclosion. MB = midbrain (central cortex), OL = optic lobe.
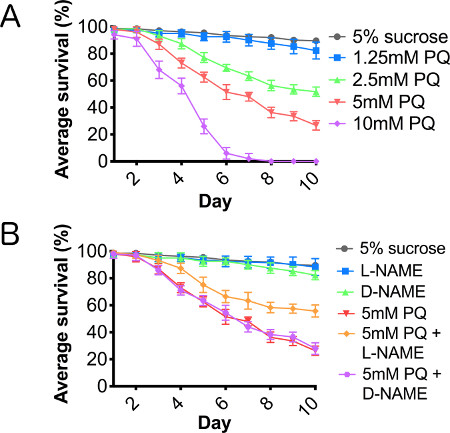
Figure 2. Paraquat results in a dose-dependent reduction in lifespan, mediated through nitric oxide synthase. Wild type male flies fed (A) 5% sucrose (control), and serial dilution of paraquat ranging from 1.25-10 mM concentration in 5% sucrose and (B) cofed paraquat with L-NAME or D-NAME for 10 consecutive days. N = 3 groups of 10. Error bars = SEM.
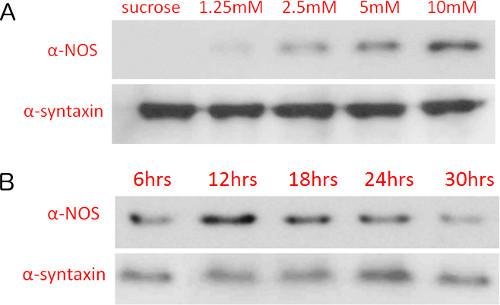
Figure 3. Nitric oxide synthase protein levels increase with paraquat concentration and length of exposure. Immunoblot detection of nitric oxide synthase levels of flies treated with varying (A) paraquat concentrations (12 hr exposure) and (B) 10 mM paraquat with varying exposure durations. Protein levels were detected on nitrocellulose membranes using chemiluminescence and X-ray film exposure.
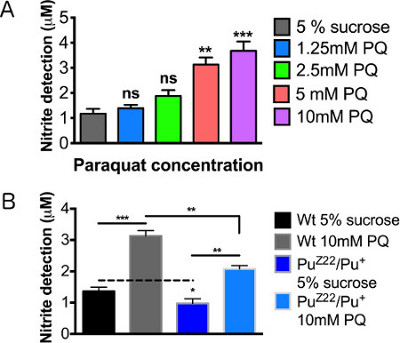
Figure 4. Nitric oxide detection levels after paraquat exposure. Male wild type (WT) flies 2-3 days posteclosion were assayed following 12 hr feeding. (A) Concentration curve of paraquat concentrations. (B) Male Punch heterozygous mutants fed sucrose control and 10mM paraquat. N = 20 brains/group. Results represent 3 replications per group. Error bars = SEM. Statistical analyses were performed using one-way ANOVA with Dunnett's Multiple Comparison posttest (*, p < 0.05, **, p < 0.01, ***, p < 0.001).
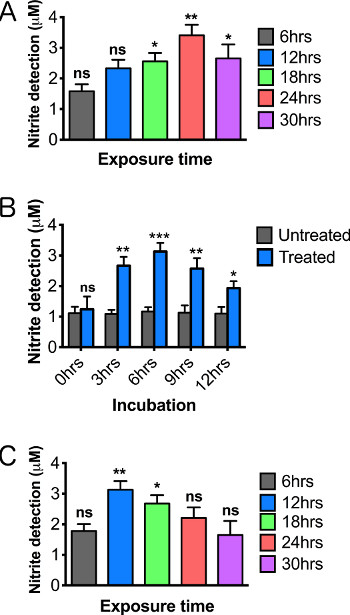
Figure 5. Optimizing the detection of nitric oxide production by altering the length of paraquat exposure and time of tissue incubation. Relative nitric oxide levels on male brains 2-3 days posteclosion following 12 hr treatment of (A) 5 mM paraquat and (B) 10 mM paraquat. (C) NO curve using varying tissue incubations periods following 12 hr, 10 mM paraquat feeding. Significance values were calculated relative to the following: (A, B) 5% sucrose controls for each exposure time (not shown in graph), (C) 5% sucrose controls for each incubation time (not shown in graph). N = 20 brains/group. Results represent 3 replications per group. Error bars = SEM. Statistical analyses were performed using one-way ANOVA with Dunnett's Multiple Comparison posttest (*, p<0.05, **, p<0.01, ***, p<0.001).
