1. Enzymatic Reaction – day 1 (1 hour in the afternoon)
- The first step is to prepare 10-20 independent enzymatic reactions in which IP6K1 or VIP1 convert IP6 to the pyrophosphorylated isoforms.
- We use His-IP6K1 and GST-Vip1 enzymes purified from E. coli according to the protocol previously described17,18.
- Prepare 50 μL reactions containing 1X reaction buffer (30 mM Hepes pH 6.8, 50 mM NaCl, 6 mM MgSO4, 1 mM DTT), 6 mM PhosphoCreatine (PCr), 25 U/mL CreatinePhosphoKinase (CPK), 5 mM ATP (Mg salt), 0.3 mM IP6, 0.05-0.1 μg His-IP6K1 or GST-Vip1. Adjust volume with MiliQ ddH2O.
- Briefly spin the reaction and incubate at 37°C overnight with rotation.
2. Polyacrylamide gel casting and loading – day 2 (4 hours in the afternoon)
- The polyacrylamide gel is prepared using 24 cm long,18 cm wide glass plates and 1.5 mm wide spacers. Usually a 16 lane or a preparative single lane comb is used.
- Prepare a mix (50 mL/gel) containing the following: 35.5% (w/v) Acrylamide:Bis-Acrylamide 19:1, 1X Tris/Borate/EDTA (TBE), 0.05% (w/v) ammonium persulfate (APS), 0.05% (w/v) Temed. Pour the mix between the pre-casted glass plates, insert the comb and let polymerize for 30-60 minutes at RT.
- Once the gel has polymerized, transfer the apparatus to the cold room and pre-run in 1X TBE for about 30-60 minutes at 200-300 Volts.
- Add 1X of OrangeG dye (10 mM Tris-HCl pH7.0, 1 mM EDTA, 30% glycerol, 0.1% OrangeG) to each reaction. Prepare a sample containing 2 nmol of IP6 to load as a standard control.
- Wash each well thoroughly with running buffer using a syringe and a 21G needle to remove any precipitate, then load the gel. Avoid loading on the side wells.
- Run the gel overnight at 450-550 Volts (7 mAmp/gel), until the OrangeG dye band is within the last 10 cm from the bottom of the gel.
3. Isolation of IP7 – day 3 (4 hours) and day 4 (6-7 hour SpeedVac drying process)
- Disassemble the gel apparatus and carefully remove one glass plate leaving the gel on the other one. Cut a small portion of the gel from just above the OrangeG dye band to the bottom containing the IP6 standard and one sample lane, as shown in Figure 1.
- Stain the cut portion of the gel with Toluidine Blue (0.1% (w/v) toluidine blue, 20% (w/v) methanol, 2% (w/v) glycerol) for a few minutes (1-3 min) or until the inositol pyrophosphate band appears. Put the glass plate previously removed back on top of the gel to prevent the unstained gel from drying.
The IP7 band should be visible since it runs slightly slower than the IP6 standard. ATP, which runs faster than IP6, should also be visible (Figure 1). Transfer the stained portion of the gel in a de-staining solution (20% (w/v) methanol) for a few minutes, wash away any excess of Toluidine Blue and reposition the gel with the unstained gel.
If visualization of higher pyrophosphorylated inositol isoforms (IP8 and IP9) is required, stain the gel with Toluidine Blue staining solution for 20 minutes at room temperature. Subsequently, wash away the Toluidine Blue with the de-staining solution for about 15 minutes.
- With a razor blade cut the IP7 band on the unstained portion of the gel using as reference the IP7 migrating position determined with the stained gel (Figure 1).
- Put the IP7 band that was cut from the gel on a 15 mL tube and add 10 mL of MilliQ ddH2O. Put tubes in rotation for 10 minutes at room temperature. Discard the liquid to remove excess of TBE and microscopic acrylamide particles.
- Subsequently, perform two dehydration-hydration cycles. Add 5 mL of 50% (w/v) methanol to the tube with the gel containing IP7 and rotate at room temperature for 2 hours. Transfer the gel slice to a new 15 mL tube containing 5 mL of MilliQ ddH2O and rotate at room temperature for 2 hours. Do not discard the methanol and MilliQ H2O from the tubes. Repeat the dehydration-hydration cycle once more by re-transferring the polyacrylamide gel in the 15 mL tubes previously used. One of the washes can be performed over night.
- To concentrate the eluted IP7, dry the 10 mL together (5 mL MilliQddH2O and 5 mL 50% (w/v) methanol) using a SpeedVac heated at 60°C.
- Once the samples are nearly dry, transfer the remaining liquid (300-600 μl) to a1.5 mL centrifuge tube and spin for 2 minutes at 5000 rpm.
- Collect supernatant and transfer into a fresh 1.5 mL centrifuge tube; leave the bottom 20-30 μL since it may contain acrylamide particles.
- If necessary continue the drying process using an unheated SpeedVac. The recovery of IP7 is dramatically reduced if the samples dry completely, therefore terminate the drying process when the samples reach the volume of 100-300 μL.
4. Determination of IP7 concentration and purity.
- Use 2-5 μL of the recovered IP7 sample to run on a PAGE gel, similarly to Sections 2.2-2.5. Load several dilutions of IP6 (i.e. 0.5, 1, 2, 4 nmol) as concentration standard and 4 nmol of Poly-P marker. After running the gel, visualize the inositol pyrophosphate isoforms by staining and de-staining the entire gel with Toluidine Blue solution, following the procedure described in Section 3.2 (Figure 2A).
- After Toluidine staining, the concentrations can be determined by scanning the gel and comparing the differences in intensity between IP6 and IP7, using imaging software such as Image-J, as shown in Figure 2B.
5. Representative Results:
The preparative enzymatic conversion of IP6 to IP7 using IP6K1 and VIP1 enzymes can be easily resolved using PAGE analysis (Figure 1). The loading of IP6 as a size control together with Toluidine Blue gel staining allows the identification of the pyrophosphorylated derivates, since they run slower depending on the number of phosphate groups present on the inositol ring. The procedure described above allows the easy purification of IP7. The analysis of the purified inositol pyrophosphate by PAGE revealed the purity of our IP7 (Figure 2A). Interestingly, the 1/3PP-IP5 isomer of IP7 product of VIP1 migrates slightly slower than the 5PP-IP5 isomer of IP7 that is generated by the IP6K1. Use of IP6 standards permit an easy quantification of the concentration of the purified IP7 (Figure 2B). Before using IP7 for further experiments, its biological activity can be assessed
(Figure 3). 5PP-IP5 is incubated with VIP1 and with the IP7 phosphatase DDP1 (diphosphoinositol polyphosphate phosphohydrolase). Routinely, the purified IP7 is converted to IP8 by VIP1 and to IP6 by DDP1 (Figure 3).
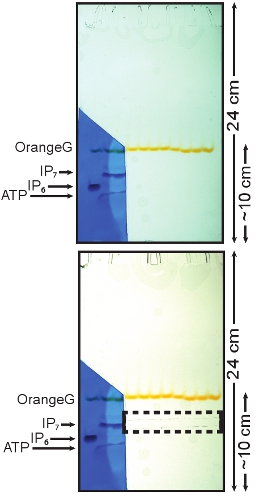
Figure 1: Toluidine staining of PAGE and isolation of the IP7 band. The portion of the gel containing the standard (IP6) was cut and stained using a Toluidine Blue solution. The three bands represent (top to bottom) IP7, IP6 and ATP. The stained portion of the gel was then aligned with the remaining of the gel. This allows the localization of the portion of the gel containing IP7, which can then be cut and purified (dashed box).
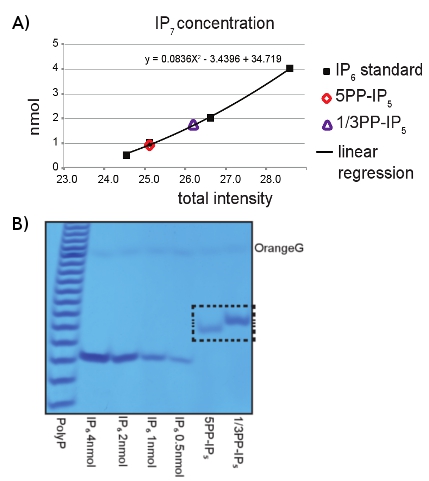
Figure 2: PAGE analysis of IP6K1 and VIP1 reaction products. A) Analysis of IP6 (4, 2, 1, 0.5 nmol) by Toluidine Blue staining was used in order to determine the IP7 concentration purified from both IP6K1 (5PP-IP5) and VIP1 (1/3PP-IP5) reactions. B) Scatter plot analysis to determine the concentration of the purified IP7. Concentrations were determined according to band intensity, calculated using imageJ software, compared to pre-determined amounts of IP6. The X-axis represents intensities; the Y-axis represents concentrations expressed in nmol.

Figure 3: Analysis of IP7 biological activity. To determine the quality of the purified IP7 we incubated 5PP-IP5 (IP6K1 generated IP7) with VIP1 or with the IP7 phosphatase DDP1 and then resolved the reaction on PAGE. The DAPI and Toluidine staining revealed the expected production of IP8 by VIP1 and the conversion of IP7 to IP6 by DDP1.
