The above described protocol details a method that allows the investigation of both early- and late-stages of organogenesis, often limited by complex cellular and molecular interactions.
This method was previously employed in Figueiredo et al.5 to unravel the role of Notch and Hh signaling in the avian parathyroid/thymus common primordium development.
Herein, new results are shown in Figure 1 and Figure 2 using the same model of organogenesis. Figure 1A depicts the experimental design used to explore the early-stages of thymus and parathyroid formation. The quail embryonic territory comprising the prospective organ rudiments (3/4PAR) was isolated and grown in vitro for 48 h in an organotypic system.
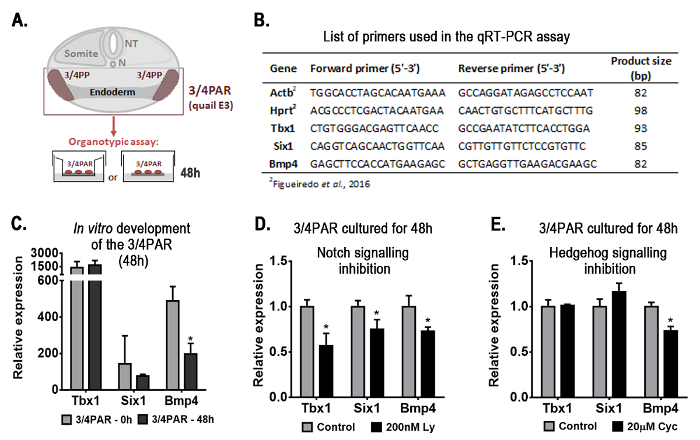
Figure 1. Representative results obtained with the organotypic culture assay: gene-expression analysis of the embryonic region containing the presumptive territories of the thymus and parathyroids (3/4PAR) developed in vitro for 48 h. Schematic representation of the transversal section of the embryo at the region of interest and the experimental design (A). Briefly, the 3/4PAR at qE3 was mechanically isolated and grown in vitro for 48 h. The expression of the 3/4PAR-related genes, Tbx1, Six1, and Bmp4, was examined by qRT-PCR using the primers in the table (B). The expression of Tbx1, Six1, and Bmp4 was analyzed in freshly isolated (3/4PAR-0 h) and cultured (3/4PAR-48 h) tissues (C). The expression of PAR-related genes was analyzed in tissues grown in vitro for 48 h in the presence of 200 nM Ly411575 (D) and 20 µM cyclopamine (E), which are pharmacological inhibitors of Notch and Hedgehog signaling pathways, respectively. Expression of each transcript was measured as a ratio against the mean of the β-actin and hypoxanthine-guaninephosphoribosyltransferase transcript expression levels and expressed in arbitrary units (each transcript in the control = 1). Means and standard deviations were determined with a software for biostatistics analysis and scientific graphic design. Error bars represent standard deviations of the mean. Two-tailed unpaired Student's t-test was used and results were considered significantly different when the p-value was less than 0.05 (p < 0.05). β-actin, Actb; cyclopamine, Cyc; Hypoxanthine-guaninephosphoribosyltransferase, Hprt; LY-411.575, Ly; N, Notocord; NT, Neural Tube; PAR, pharyngeal arch region; PP, pharyngeal pouch. Please click here to view a larger version of this figure.
The expression of genes known to be involved in the formation of PAR structures (PAR-related genes), i.e., Tbx116,17, Six118, and Bmp415,17, was evaluated during the normal development. Quantitative real time PCR (qRT-PCR) was performed as previously described5 (primers are listed in Figure 1B). Transcripts of the three genes were detected in freshly isolated (3/4PAR-0 h) and in 48 h-cultured tissues (3/4PAR-48 h) (Figure 1C). Only Bmp4 expression levels were significantly decreased after 48 h of culture.
To evaluate the role of Notch and Hh signaling pathways in the early-stages of thymus and parathyroid development, pharmacological inhibitors were added to the culture medium during in vitro development. Inhibitor doses are described in Figueiredo et al.5 The expression levels of the three genes analyzed were significantly reduced in the 3/4PAR grown in the presence of Notch inhibitor, when compared to control conditions (without drug) (Figure 1D). Conversely, only Bmp4 transcripts were significantly reduced in the 48 h-cultured tissues when Hg signaling was blocked (Figure 1E).
To study the late-stages of thymus and parathyroid gland organogenesis, cultured tissues were then grafted onto CAMs and allowed to further develop for 10 days (see the experimental design in Figure 2A).
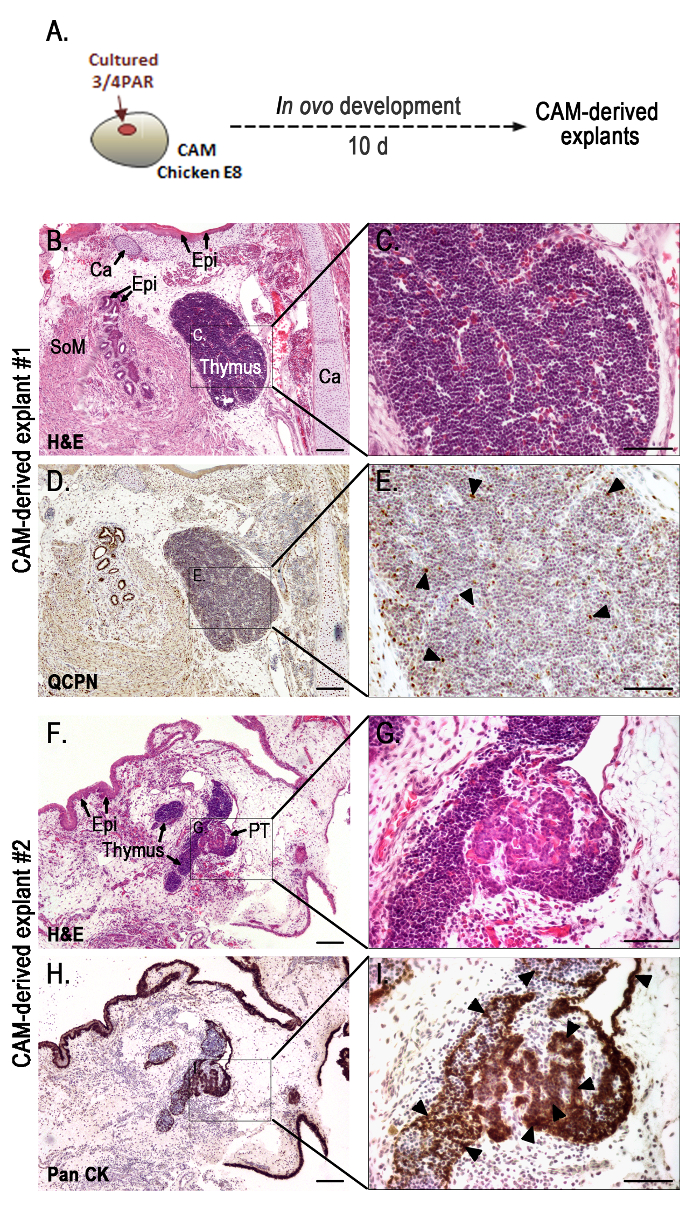
Figure 2. Representative results obtained with the in ovo assay: morphological analysis of the grafts grown for 10 days in the chorioallantoic membrane. Schematic representation of 48 h-cultured PAR grafted onto the CAM and developed for 10 days (A). Serial sections of CAM-derived explants (B–I) slides stained with H&E (B, C, F, and G), immunodetected with QCPN (D and E) and anti-Pan CK (H and I) antibodies, and counterstained with Gill's hematoxylin. Black arrow heads indicate strong immunostaining for QCPN (E) and Pan CK (I). A transverse section of a chimeric thymus with lymphoid cells of host origin and quail-derived thymic epithelial cells with strong QCPN+ signals (black arrowheads) (E). Strong Pan CK+ signals (black arrowheads) in the epithelia of the thymus and parathyroid glands (I). Images were collected using imaging software and a microscope with a camera (see Table of Materials). Ca, cartilage; CAM, chorioallantoic membrane; Epi, epithelium; PAR, pharyngeal arch region; PT, parathyroid glands; SoM, smooth muscle; 10 d, ten days. Scale bars, 50 µm (B, D, F, and H) and 100 µm (C, E, G, and I). Please click here to view a larger version of this figure.
Morphological analysis of organs developing on CAM-derived explants was performed by conventional histology and immunohistochemistry (Figure 2B–I), as previously described5. CAM-derived explants showed fully formed chimeric thymus (Figure 2B–E) with quail-derived (QCPN+) thymic epithelium colonized by lymphoid progenitor cells of donor origin (chicken) (Figures 2D, E). Serial sections of CAM-derived explants further processed for immunocytochemistry with anti-pan cytokeratin (anti-pan CK) antibody (an epithelial cell marker), showed thymic and parathyroid epithelia with normal morphological features (Figure 2H, I). The thymic epithelial cells displayed a reticular architecture while parathyroid parenchymal cells were globular, arranged in clusters and encircled by numerous capillaries. Additionally, other PAR-derived structures from the respiratory apparatus could be observed in the grafts. Cartilage, respiratory epithelium, and smooth muscle associated to the mucosa were easily distinguished in Figure 2B.
