All animal experiments were conducted in compliance with the Animal Welfare Act, the implementing Animal Welfare Regulations, and the principles of the Guide for the Care and Use of Laboratory Animals. Prior testing has demonstrated that these methods are reliable26 and have no adverse effects on the health or limb function of the pig. Testing has been conducted as often as weekly without any adverse events23. Additionally, testing pre- and post-surgical interventions during the same day can be performed without placing untoward stress on the animal or inducing neuromuscular dysfunction.
1. Computer set up
- Ensure initial set and calibration of the apparatus and components are conducted under manufacture specifications (see Table of Materials). Calibration using a range of weights from 0.2-2.5 kg is suggested.
NOTE: Torque is measured by a 140 mm foot-pedal (0.14 m) attached to a linear torque sensor with a 50 Newton-meter (N·m) capacity. The instrument's gain is set to scale to 25 N·m capacity by default to better match the anticipated torque production. Calibration is performed by applying a known mass (e.g., 1 kg) to the foot-pedal at a known distance (e.g., 100 mm from the axis of rotation) and calculating torque. For example, 1 kg equals 9.806 N applied at 0.1 m is 0.9806 N·m torque. A relationship can then be established between torque applied to the torque sensor and the corresponding voltage output by the torque sensor. The author's torque sensors have confirmed the linearity of this relationship from 0.2-20 kg applied to a particular 40 cm calibration plate. Due to the length of the standard pedal, a calibration range of 0.2-2.5 kg is recommended. This produces enough signal to calculate the scale factor by linear regression. - Turn on the computer, stimulator, transducer system, and the analog-digital interface about 30 min before testing to allow stabilization of heat-related material changes that can impact electrical properties. Select the appropriate and connected data acquisition (DAQ) device.
- Set up experimental parameters in the software as needed; the software allows for a saved study template. Prepare to set up the experiment (i.e., study template) to create a new study using the Create a New Study workbook option.
NOTE: Experimental parameters can be pre-loaded before beginning the study, which will result in prompts to include additional specific experiment information such as sex, body mass, date of birth, the time point of testing, treatment group, or similar variables as needed. The study setup parameters can be saved and used across the experiment. - Select the study previously created at the beginning of each evaluation. Add a New Animal if this is the first test for the pig to be tested and follow the prompt for the variables input into the study.
- Click on Prepare Experiment once ready to begin the study, which will be needed to optimize electrode placement. Deliver repeat twitches to the nerve while determining optimal placement once the electrodes are placed (see step 3.6.).
- Click on Configure Instant Stim first, and then adjust the pulse frequency, pulse width, number of pulses, train frequency, and run time.
- Then, click on Instant Sim to deliver repeat twitches. Alternatively, push the Manual Trigger button on the Stimulator unit to manually give one twitch.
- Open the Live Data Monitor during the study protocol when ready to start the whole experiment to allow real-time investigation/visualization of the contractions. Click on Run Experiment when prepared to begin the experiment (following animal preparation, see step 2).
2. Anesthesia preparation and maintenance
- Fast male or female pigs, 40-90 kg, overnight before anesthesia event, allow water ad libitum. Obtain and record the correct bodyweight of the pig on the day of the procedure.
- Induce anesthesia with intramuscular injections of tiletamine/zolzepam (Telazol, 4-6 mg/kg), xylazine (1-3 mg/kg), and propofol (2.6 mg/kg). Initially maintain with 5% isoflurane via facemask.
- Intubate the pig with an endotracheal tube and place it on an automatic ventilator. Maintain the pig on peak pressure at 20 cm H2O, an initial tidal volume of 10 mL/kg, and respiration rates at 8-12 breaths/min.
- Adjust the ventilator setting to maintain an end-tidal PCO2 of 40 ± 5 mmHg. Maintain anesthesia with 1%-3% isoflurane in 30%-37% O2.
- Maintain the body temperature of the pig at 37 °C for the duration of the protocol. Insert ear vein and Foley catheters for fluid delivery and urine collection, as needed.
NOTE: Using surgical plane anesthesia will prevent secondary contractions during the testing, especially from the gluteal muscles. - Monitor the depth of the anesthesia via eye reflex and position, lack of jaw tone, heart rate (range 80-150 bpm), systolic blood pressure (range 120-70 mmHg), or a combination of all these signs.
- Prepare both the right and left hindlimbs once the pig is fully anesthetized and stable by first cleaning the limbs with soap and water to remove any debris and then shave the hair from the skin. Pay close attention to the lateral knee area, which will be used for electrode placement later.
- Transport the pig to a surgical table and securely place it in the supine position. Position the pig toward the foot of the table with the gluteal muscles at or slightly over the end of the table.
NOTE: This will allow the surgical table and table holding the testing apparatus to abut. - Extubate the pig after the test and allow them to recover. Standard pig food and water should be replaced once the pig is fully recovered and can ambulate freely within the cage.
NOTE: Post-procedure analgesia is unnecessary for the in vivo testing alone; however, carprofen and/or buprenorphine SR can be provided per veterinary recommendation. Consultation with a local veterinarian is encouraged. The anesthesia and medications listed here are for guidance only and are currently approved at the University of Minnesota. Maintenance of anesthesia with isoflurane was chosen based on its rapid onset and offset and its minimal impact on in vivo nerve stimulation evoked torque29. Take care to have consistency in anesthesia parameters across studies. During the protocol, anesthesia assessment and recording is conducted at 15 min intervals; recording is undertaken based on local Institutional Animal Care and Use Committee (IACUC) and United States Department of Agriculture (USDA) guidelines and requirements.
3. Evaluation of in vivo isometric torque
- Place the foot on the foot-plate of the force transducer. Use a flexible cohesive bandage to attach the foot to the foot-plate.
NOTE: An entire role per foot is necessary; ideally, the 4-inch x 5-yard role is adequate. - Hold the foot in position on the foot-plate with the ankle at the neutral (A) and secure the foot to the plate by wrapping the cohesive bandage around the foot and foot-plate in the style of a closed basket weave ankle taping (B).
NOTE: The two study personnel will be required to simultaneously perform the individual (A) and (B) tasks. - Position the ankle at a right angle once the foot is secured to the foot-plate, defined as 0° or neutral for reference of degrees of plantar or dorsiflexion.
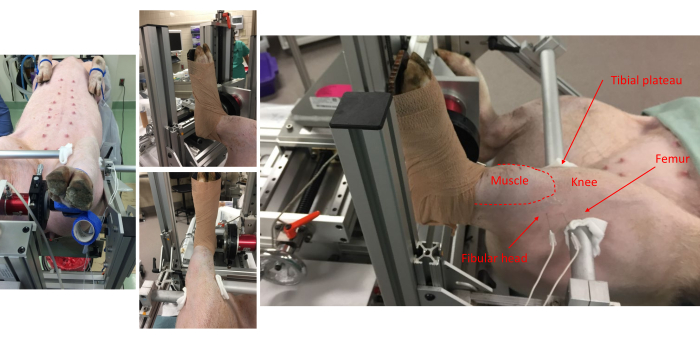
Figure 1: Pictures from various vantage points show the pig attachment to the foot-plate and anatomical alignment onto the frame. Anatomical landmarks for the anterior compartment muscles, fibular head, knee, tibial plateau, and femur are noted. Note the placement of subdermal electrode pairs on the lateral side of the leg. Please click here to view a larger version of this figure.
- Stabilize the knee and ankle at right angles.
- First, position the limb clamping bars close to the needed locations. When ready, starting on the medial aspect of the limb, align the limb clamping bar at about the tibial plateau.
- Then, align the lateral limb clamping bar at the distal head of the femur.
NOTE: Between the end of each limb, the clamping bar uses a folded 4 x 4 gauze pad to protect the skin adjacent to the bar. - Stabilize the bars tightly using the locking thumbscrews.
NOTE: The limb clamping bars will not be in line with each other but will align with the pig's anatomy. - Clean the skin around the fibular head by applying 70% alcohol via clean gauze in concentric circles starting at the center of intended electrode placement and moving outwards. Place the sterile percutaneous needle (50 mm, 26 G monopolar) and electromyography (EMG)-style electrodes (see Table of Materials) across the peroneal nerve. Implant electrodes subdermally, approximately 5-10 mm.
- Optimize the electrode placement using increasing current amplitudes, as adjusted on the stimulator. Start at 100 mA and increase as needed.
NOTE: 300-500 mA is usually required for peak twitch torque. - Visualize the twitch torque magnitude on the live data view and over the pig's anterior compartment; the hooves may splay and move upward as well.
- Ensure that the posterior compartment, or tibia nerve, is not activated during stimulation. Visually inspect and palpate posterior compartment contraction and downward movement of hooves during stimulation.
- Inspect the plateau region of tetanic contraction from the live torque-time tracing in the following steps for lack of antagonist muscle recruitment (i.e., plantarflexion for this protocol).
- Elicit maximal isometric tetanic torque using the following stimulation parameters: 100 Hz, 0.1 ms pulse width, over an 800 ms train17, once the electrode placement and stimulation amplitudes are optimized.
NOTE: These parameters can be used for various contractile evaluations.
4. Protocol for torque-joint angle analysis
- Measure the maximal isometric tetanic torque across a range of ankle positions ranging from neutral to the near end ranges of plantarflexion, or 0-50° of plantarflexion.
NOTE: Using 10° increments will require six contractions, and the incremental change can be adjusted for specific experimental questions. - Start loosening both locking screws of the goniometer stage to move between joint angles. Ensure both locking screws are tightened before the next contraction.
NOTE: The goniometer is scribed with degree markings to allow for precise alignment. It is likely 0° of plantar flexion, which is offset by 180° on the goniometer. Take caution to ensure intended positioning. - Determine the time between the contractions experimentally; however, 2 min is sufficient to avoid fatigue.
NOTE: As the ankle joint angle is incrementally changed, the needle electrodes may shift. It may be necessary to confirm the placement of the electrodes with twitch contractions, as noted above (see step 3.8).
5. Protocol for torque-frequency analysis
- Position the ankle at the desired joint angle. Take care, experimentally, to conduct testing at the same joint angle each time.
NOTE: Typically, torque-frequency analyses are performed at a single joint angle corresponding with peak isometric torque derived from the torque-joint angle analysis. Peak torque is produced at ~30-35° of plantarflexion. - Measure maximal isometric torque over a range of stimulation frequencies that induce unfused trains of twitches up to and beyond those that induce fully fused tetani.
NOTE: This can be achieved by stimulating at 10, 20, 40, 60, and 100 Hz (0.1 ms pulse width; 800 ms train) with 2 min between each contraction to avoid fatigue. Depending on exact experimental questions and specific pig models, frequencies may be adapted. The bioenergetic substrate most likely utilized during an 800 ms contraction to maintain intracellular ATP is phosphocreatine30, and the resynthesis of phosphocreatine relies on the phosphocreatine shuttle31. Phosphocreatine recovery kinetics indicate a 90% or more remarkable recovery between 90-120 s after contraction ends30. Therefore, the recommended rest intervals between contractions are 90-120 s. Although, this may be influenced by experimental designs, including muscle disease, injury, and/or aging.
6. Data analysis
- Click on Analyze Results if still in the software to open the Analysis window. Alternatively, open the Analysis program directly.
- Whether using an automated data platform or manual analysis, calculate the different variables in analyzing individual isometric waveforms.
NOTE: These variables include: maximal twitch torque, maximal tetanic torque, and contractile properties related to twitches and tetani, e.g., time-to-peak and half-relaxation. Many experimental variables can normalize force, for example, body weight, muscle volume determined from MRI (magnetic resonance imaging) or CT (computed tomography), or terminal muscle weight. Both absolute torque (N·m) and torque normalized to body mass (N·m/kg) are presented. The resting torque placed on the foot-plate will differ across experiments. A baseline correction for resting torque should be applied to ensure true maximal twitch and tetanic torques are recorded. Baseline torque at each joint angle is recorded and can indicate changes in passive torque.
Reliability and optimization of the in vivo testing parameters of the anterior compartment of the pig have previously been reported26. Comparative data across rodents and pigs for torque-frequency has also been reported27.
During the in vivo assessment, visualization of the torque waveform is needed in real-time to ensure appropriate anterior compartment activation. The waveforms should only reflect dorsiflexion. The waveforms should have a smooth, rounded appearance and an apparent tetanic plateau (Figure 2A). Inconsistencies or perturbations of the waveform indicate various experimental limitations, such as inadequate stimulation, improper electrode placement, or inadequate depth of anesthesia (Figure 2B).
Figure 3A is a twitch torque-time tracing with an arrow indicating 50% max torque. Time-to-peak contraction should start at the initiation of the stimulator and end when maximal twitch torque is achieved (representative time bars are shown below the tracing). Half-relaxation for a twitch should start at the maximal twitch torque and end at 50% maximal twitch torque (representative time bars are shown below the tracing). Figure 3B is a tetanic torque-time tracing with an arrow indicating 50% max torque. Unlike twitches that are ideal in terms of a definitive and timely maximal torque, tetanic contractions have greater variability in the timing of maximal torque concerning when the stimulator starts and ends, requiring a more nuanced approach to contractile property analysis. Time-to-peak contraction should begin with the initiation of the stimulator and stop somewhere between 90%-100% of maximal torque. The time bars in Figure 3B show a cutoff of 95% maximal torque. This is helpful in cases such as the selected representative data because the maximal torque is not reached until late in the plateau phase. A complementary analysis to time-to-peak is the average rate of contraction. The dashed bars on the ascending limb of the torque-time tracing represent a range of 30%-70% maximal torque. The average rate of contraction should be started at the start of stimulation and capture the average rate change between 30%-70% maximal torque. These are recommended ranges, and individual research groups can determine the ideal range around 50% (e.g., ±10%). The important part is to be consistent within and across studies. In contrast to the twitch, tetanic contraction half-relaxation should start at the end of stimulation instead of maximal torque for the same reason mentioned above with time-to-peak. The time bars in Figure 3B represent the time between the end of the stimulation and reaching 50% relaxation. A complementary analysis to half-relaxation is the average rate of relaxation. The dashed bars on the descending limb of the torque-tracing represent the same 30%-70% maximal torque range as the ascending limb. The average relaxation rate should start at the end of stimulation and capture the average rate of change between 30%-70% maximal torque. Again, these are recommended ranges. One critical note: do not confuse the average rate of contraction/relaxation with the maximal rate of contraction/relaxation. The maximal rate represents the single most remarkable rate change between two adjacent data points and can be widely variable.
Several twitch and contractile properties can be analyzed to gain insight into fiber type and excitation-contraction coupling attributes of the skeletal muscles10,32. Over-interpretation of the twitch and contractile properties are cautioned; they represent suggestions and rationale for further cellular-level interrogation and are not necessarily indicative. In general, rates of contractility can reflect sarcoplasmic reticulum calcium release and myosin heavy chain isoform enzymatic rate. In contrast, relaxation rates can reflect sarco(endo)plasmic reticulum calcium ATPase enzyme rate and isoform. These properties can be influenced by fatigue, muscle damage, exercise training, and numerous pathologies (e.g., disuse atrophy).
Figure 4 depicts representative values for torque-frequency and torque-joint angle relationships for uninjured limbs. These data are representative of a wide range of pig sizes.
A representative, experimental analysis of surface EMG was conducted during in vivo muscle analysis (Figure 5) to demonstrate experimental control of rate coding and total muscle activity. Adhesive EMG electrodes were placed at the mid-belly of the peroneus tertius. A ground electrode was placed on the knee to minimize stimulation artifact, and stimulation electrode needles were placed around the peroneal nerve proximal to the muscle location. Simultaneous torque and EMG recordings were made at stimulation frequencies of 20, 60, and 100 Hz. The number of stimulator pulses (red bars in Figure 5) reflects the quotient of stimulation duration and time between pulses. For example, a 20 Hz stimulation frequency means a pulse every 50 ms; therefore, 400 ms stimulation duration divided by 50 ms between pulses equals eight pulses delivered (Figure 5A). The stimulator pulses are delivered to the nerve axon via percutaneous needle electrode placement and produce a similar number of electrical muscle pulses (i.e., 20 Hz equals 8 EMG recordings), demonstrating the experimental control of action potential frequency of the muscle group of interest. The raw EMG recordings can be converted via root-mean-square analysis (EMG RMS) to visualize the total muscle activity with increasing stimulation frequency. The area under the curve (AUC) analysis is one way to quantify the EMG RMS to determine changes in whole muscle activity. Representative AUCs for each EMG RMS stimulation frequency is provided (Figure 5A–C).

Figure 2: Representative high and low-quality waveforms. (A) Isometric waveforms present in a square-wave appearance, with a notable fluid plateau. (B) Low-quality waveforms can be due to inadequate stimulation or improper electrode placement. In these instances, repositioning of the electrodes is needed. For both A and B, the stimulator pulses (red bars) are indicated. Please click here to view a larger version of this figure.
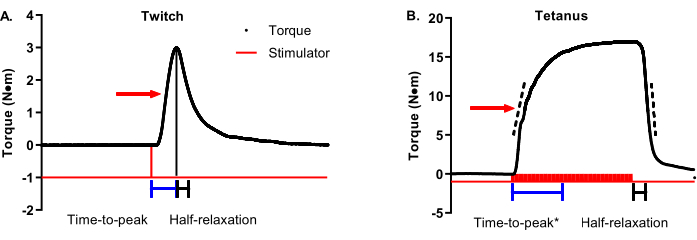
Figure 3: Twitch and tetanic contractile property analysis. (A) Representative twitch (1 Hz) and (B) tetanic (100 Hz) torque-time tracings are modified to detail contractile properties. The red arrow on each graph shows 50% maximal torque. Blue and black bars beneath the tracings show time-to-peak and half-relaxation time durations, respectively. Dashed bars on the ascending and descending limbs of the tetanic torque-time tracing represent a range of 30%-70% maximal torque that can be used to determine the average rate of contraction or relaxation. Please click here to view a larger version of this figure.
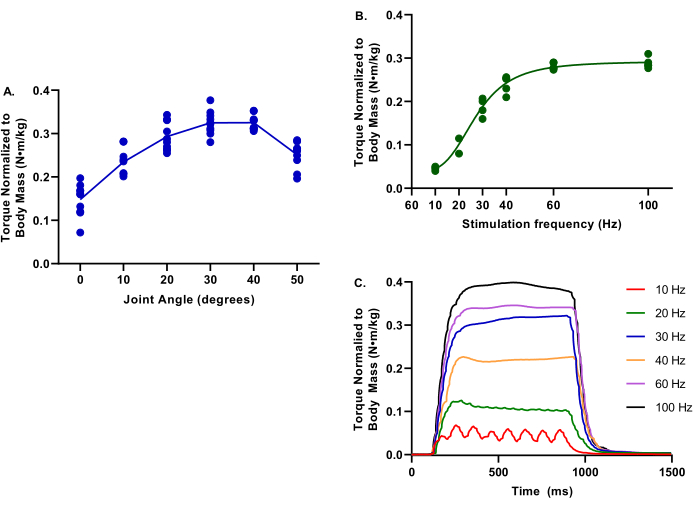
Figure 4: Example data of torque-joint angle and torque-frequency. Data provided is from a range of female Yorkshire Cross pigs at 2.9-6.3 months; 39.4-75.4 kg body mass; all considered healthy control at the evaluation time. During all testing, core body temperature was maintained at 37 °C. (A) Torque normalized to body mass is evaluated at ankle joints of 0-50° of plantarflexion; note the peak torque is at determined at 30°. (B) Torque normalized to body mass is evaluated at various stimulation frequencies from 10-100 Hz; note these evaluations were conducted with the ankle joint at 30° of plantarflexion. (C) Individual torque tracings for each of the stimulation frequencies were evaluated. Please click here to view a larger version of this figure.
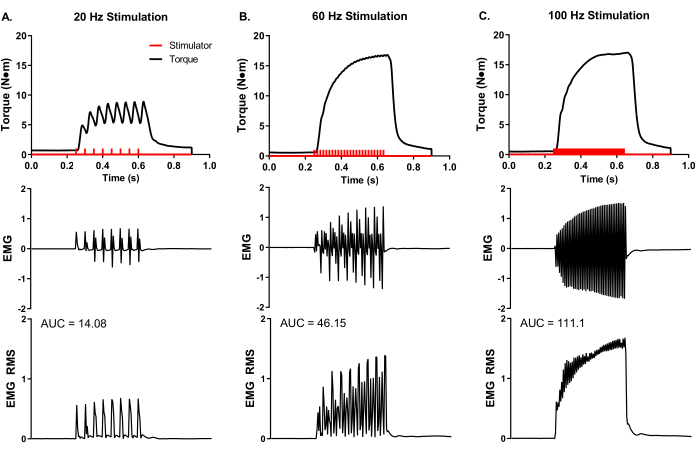
Figure 5: Concurrent in vivo isometric torque and EMG measurements. Simultaneous EMG and torque recordings at representative stimulation frequencies of (A) 20, (B) 60, and (C) 100 Hz collected from a female Yorkshire pig (~90 kg body mass). Stimulator pulses (red bars) were delivered according to the set stimulation frequency. Raw EMG recordings were converted to root-mean-square (EMG RMS) to visualize total muscle activity with increasing stimulation frequency. Representative EMG RMS curves were analyzed for the area under the curve (AUC), and AUCs are provided for each stimulation frequency. Please click here to view a larger version of this figure.
