Initially, the phenotype and purity of CB-SC were examined by flow cytometry with CB-SC-associated markers such as leukocyte common antigen CD45, ES cell-specific transcription factors OCT3/4, and SOX2. CB-SC display high levels of CD45, OCT3/4, SOX2, CD270, and galectin 9 expression, but no expression of CD34 (Figure 1A). Flow cytometry analysis confirmed the expression of exosome-specific markers including CD9, CD81, and CD63 were on CB-SC-derived exosomes (Figure 1B). Morphology and size distribution of exosomes were characterized by TEM and DLS (Figure 1C,D), with the size of 79.38 ± 20.07 nm. Western blot further proved the expression of the exosome-associated marker Alix, without expression of the ER-associated marker Calnexin (Figure 1E).
PBMC were treated with Dio-labeled CB-SC-Exo. The microscopy observation demonstrated the direct interaction of Dio-labeled CB-SC-Exo with PBMC (Figure 2A). To better define which cell population interacted with the Dio-labeled CB-SC-Exo, different cell compartments were gated with cell-specific markers such as CD3 for T cells, CD11c for myeloid dendritic cells (DC), CD14 for monocytes, CD19 for B cells, and CD56 for NK cells (Figure 2B). After an incubation for 4 hr, flow cytometry demonstrated that different blood cell compartments displayed at different median fluorescence intensity (MFI) of Dio-positive exosomes (Figure 2C). Notably, monocytes exhibited higher median fluorescence intensity of Dio-positive CB-SC-Exo than those of other immune cells (Figure 2C), highlighting that monocytes were primarily targeted by the CB-SC-derived exosomes.
To explore the direct effects of CB-SC-derived exosomes on monocytes, the purified CD14+ monocytes were co-cultured with CB-SC-derived exosomes for 3 days. The exosome-treated monocyte successfully differentiated into spindle-like morphologies (Figure 3A). Next, phenotypes of the CB-SC-Exo treated or untreated monocytes were tested, revealing the expressions of M2-associated markers including CD163, CD206, CD209 were markedly increased among the exosome-treated group (Figure 3B, red histogram). Comparing with the conventional M2 macrophages generated by M-CSF + IL-4, CB-SC-Exo-treated monocytes expressed similar levels of M2-associated markers such as CD163, CD206, CD209, with no significant differences (Figure 3C). Therefore, the data indicates that monocytes differentiate into macrophages with M2 phenotype after the treatment with CB-SC-derived exosomes.
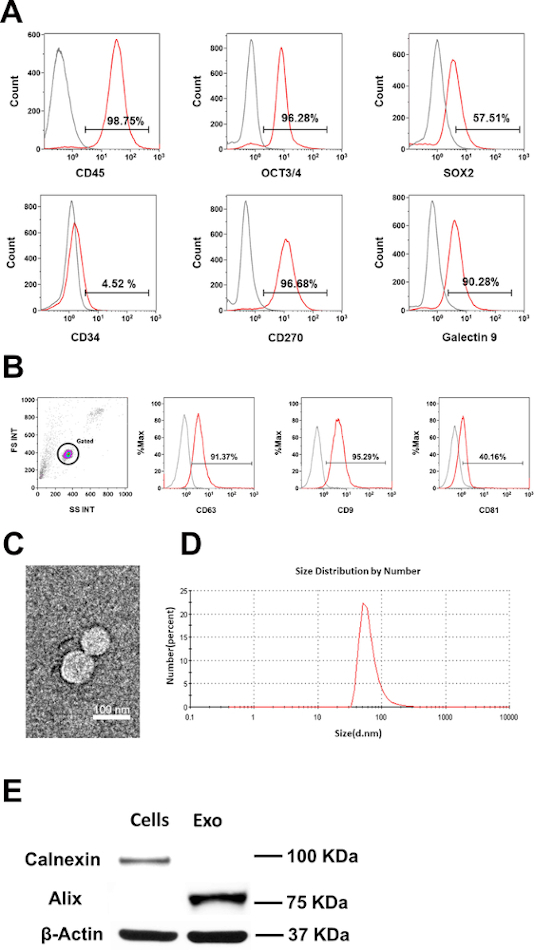
Figure 1: Characterization of CB-SC-derived exosomes. (A) Phenotypic characterization of CB-SC, high expression of CD45, OCT3/4, SOX2, CD270 and Galectin-9, no expression of CD34. (B) Expressions of exosome-associated markers (CD63, CD9, CD81) on CB-SC-derived exosomes. Isotype-matched IgGs served as controls for flow cytometry (gray histogram). (C) Transmission Electron Microscopy (TEM) image of the CB-SC-derived exosomes. (D) Size distribution of CB-SC-derived exosomes using Dynamitic Light Scattering (DLS). (E) Western blots show that CB-SC-derived exosomes display the exosome-specific marker Alix, but negative for endoplasmic reticulum (ER)-associated marker Calnexin. Please click here to view a larger version of this figure.
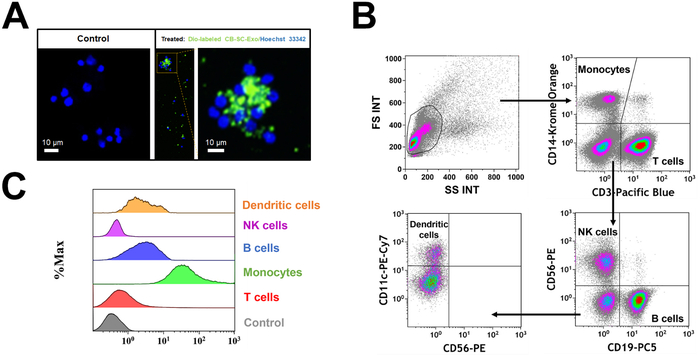
Figure 2: Interaction of CB-SC-derived exosomes with different populations of PBMC. (A) The interaction of Dio-labeled CB-SC-Exo (green) with PBMC (blue, nuclear staining with Hoechst 33342) was photographed with Nikon Eclipse Ti2 microscope with NIS-Elements software version 5.11.02, with a high magnification showing the distribution of Dio-labeled exosomes (green) in the PBMC cells after the co-incubation for 4 h 5% CO2 in the non-tissue culture-treated 24-well plate. n = 2. (B) Gating strategy for flow cytometry analysis with cell-specific surface markers for different subpopulations in PBMC, including CD3 for T cells, CD14 for monocytes, CD19 for B cells, CD56 for NK cells, and CD11c for DCs. (C) Display different median fluorescence intensity (MFI) of Dio-labeled exosome among different PBMC subpopulations (e.g., T cells, Monocytes, B cells, NK cells, DCs). Please click here to view a larger version of this figure.
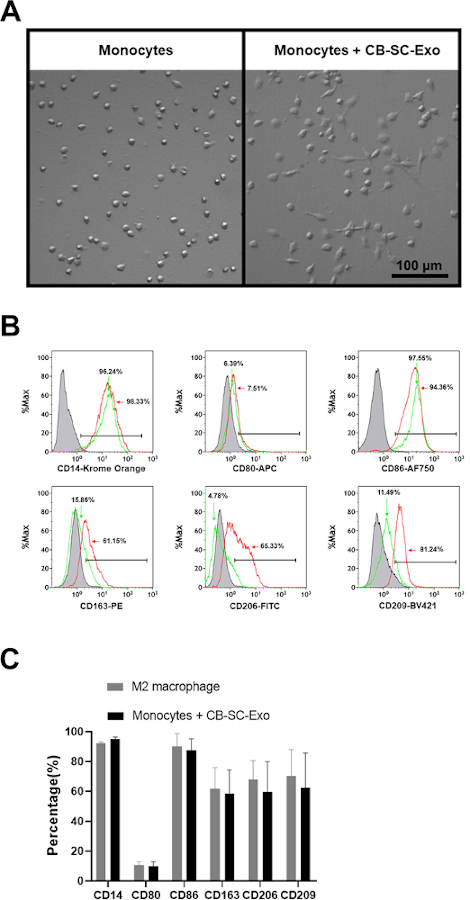
Figure 3: Effects of CB-SC-derived exosomes on monocytes. (A) Morphological change of monocytes into the spindle-like cells after treatment with CB-SC-derived exosomes. (B) Up-regulated the level of M2-associate markers’ expression after the treatment with CB-SC-derived exosomes, such as CD163, CD206, and CD209 (red line). Untreated monocytes (green line) served as control. Isotype-matched IgGs served as negative controls (gray line). (C) Phenotypic comparison between conventional M2 macrophages and the CB-SC-Exo-induced M2 macrophages. To generate the conventional M2 macrophages, the purified CD14+ monocytes were treated with 50 ng/mL macrophage colony-stimulating factor (M-CSF) at 37 °C, 5% CO2 conditions for 7 days, and followed by the overnight treatment with 10 ng/mL IL-4. M2-associated markers including CD14. CD80, CD86, CD163, CD206, and CD209 were evaluated by flow cytometry. Isotype-matched immunoglobulin G (IgG) serve as control. The data is presented as mean ± SD; N = 3. Please click here to view a larger version of this figure.
