Determining the optimal number of PECs for ex vivo mast cell degranulation assay
Mast cells (c-kit+·IgE+ double positive cells)15 represent only about 2% of PECs (Figure 1A). Estimating the maximum levels of mast cell-specific molecules to be detected in the culture supernatants on the assumption that 100% of the granules were released by mast cells in PECs, we measured the amounts of β-hexosaminidase16 and histamine17 in the total cell lysates prepared with different numbers of PECs: 2.5 x 105, 5 x 105 and 1 x 106 PECs, respectively. As shown in Figure 1B, significant levels of both β-hexosaminidase and histamine were detected even when the lysate was prepared with 2.5 x 105 PECs, and their levels increased proportionally as the number of PECs used for the preparation of the cell lysate increased.
Bearing those results in mind, we decided to use 5 x 105 PECs in the ex vivo mast cell degranulation assay for the following reasons. Considering that only a little over 3 x 106 PECs were isolated from one mouse (8 weeks old) and that they were to be plated evenly in 4 separate wells in a 96-well microplate (2 wells for β-hexosaminidase and another two wells for histamine assays, respectively), plating 1 x 106 PECs per well would likely cause a shortage of the cells. In addition, the results shown above indicated that a sufficient number of mast cells was in 5 x 105 crude PECs for carrying out the degranulation assay. That is, according to those results, it was expected that even in the case that only about 30 to 40% of the granules was released by mast cells, the levels of β-hexosaminidase and histamine detected in the culture supernatants after the degranulation assay would be high enough for being accurately quantified.
Next, we tried to confirm that β-hexosaminidase and histamine detected in the lysates of PECs were derived exclusively from mast cells in PECs. To do that, we depleted mast cells from PECs using a magnetic cell purification column (refer to step 1.2). Expectedly, unlabeled PECs that flew through the column were completely devoid of mast cells (Figure 1C). Also, expectedly, neither β-hexosaminidase nor histamine were detected in the cell lysate prepared with mast cell-depleted PECs (Figure 1D).
Ex vivo mast cell degranulation assay using crude PECs
We next examined the actual levels of β-hexosaminidase and histamine released by mast cells in PECs during culture with a specific antigen of IgE on their surface. To do that, we isolated PECs from the mice injected with anti-DNP-IgE and culture them (5 x 105) with DNP-BSA (0.5 ng/mL). As shown in Figure 2A, a significant level of β-hexosaminidase was begun to be detected in the culture supernatant within 5 min of culture and increased continually as the culture period was prolonged. Nevertheless, the rate of increase was diminished quickly after 10 min of culture, and the extent of degranulation gradually reached the plateau at around 50% after 30 to 40 min. Similar results were also obtained when the levels of histamine released by mast cells during the culture were examined (Figure 2B). Based on those results, we decided to culture PECs with DNP-BSA for 10 min in the following assays.
Verifying the in vivo effects of KET and DEX on mast cell with the ex vivo degranulation assay.
Ketotifen (KET) is an allergy medication with antihistaminergic activity. Different from other conventional antihistamines, however, it is known as a dual-acting antihistamine that also has a mast cell stabilizing activity in addition to the antihistaminergic activity. The mast cell stabilizing activity of KET has been explored21,22; however, mostly in in vitro studies and the studies to show how it has effect on mast cell in vivo are scarce. Dexamethasone (DEX) is another type of allergy medication23. DEX is known to impair functional activities of various types of immune cells to suppress a broad spectrum of immune responses24. To verify the effect of KET and DEX on the activity of mast cells in vivo, we were to carry out the ex vivo mast cell degranulation assay with PECs isolated from the mice treated with either of them.
Prior to the ex vivo mast cell degranulation assay, we first examined the effects of DEX and KET on anaphylactic reactions elicited by passively transferred DNP-specific IgE and DNP-BSA in PSA model. Expectedly, treatment with either KET or DEX resulted in improvement in anaphylactic symptoms in a dose-dependent manner (Supplemental Figure 1), indicating that the functional activity of mast cells was compromised by their treatments.
We also examined the numbers of PECs isolated from the mice treated with either compounds and the ratio of mast cells in PECs (Figure 3A). Treatments with KET, regardless of the doses used in the treatments, resulted in no noticeable change in the numbers of PECS isolated from the mice. In contrast, the numbers of PECs isolated from the DEX-treated mice decreased significantly when treated at 4.5 mg/kg dose, indicating that DEX treatment had effect on the viability of peritoneal cells. Still, it must be noted that the ratio of mast cells remained constant at around 2% regardless of the drugs and doses used for the treatment (Figure 3A). We also examined the effects of DEX and KET on the levels of β-hexosaminidase and histamine expressed by mast cells in the peritoneal cavity by measuring their amounts in the total cell lysates prepared with the same numbers of crude PECs (Figure 3B). Of note, the levels of both β-hexosaminidase and histamine were found to be augmented after treatment with DEX at the dose of 4.5 mg/kg. KET treatments resulted in little change in those levels.
Next, we performed the ex vivo mast cell degranulation assay (Figure 4). First, we adjusted the cell density of PECs evenly to 5.5 x 106 cells/mL to ensure that the same number of mast cells were used in the assay. PECs (90 μL) were then plated in a 96-well microplate and incubated with DNP-BSA for 10 min. When PECs from the mice treated with the high dose of either DEX or KET were incubated with DNP-BSA, the levels of β-hexosaminidase and histamine detected in the culture supernatants were found to be lowered significantly compared to those detected after incubation of PECs from sham-treated mice with DNP-BSA (Figure 4). An inverse correlation was also apparent between the levels of those molecules detected in the culture supernatants and the doses of DEX and KET used for treatments of the mice. Thus, the higher the dose used for the treatment was, the lower the levels of β-hexosaminidase and histamine detected in the culture supernatants after the incubation were.
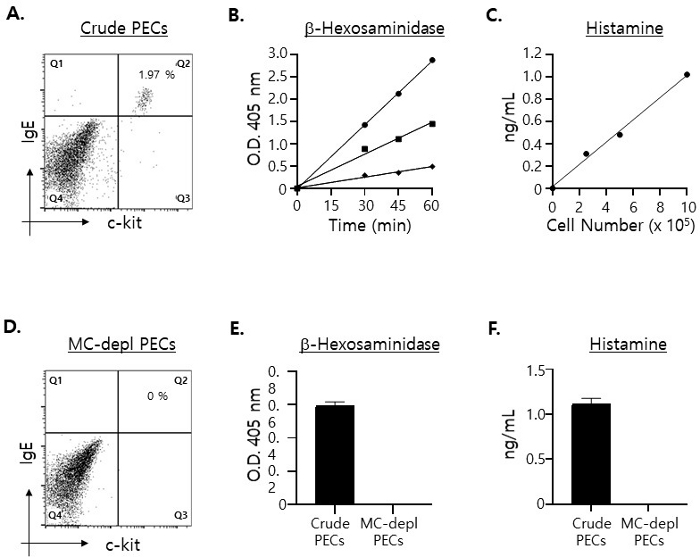
Figure 1: Quantifying the levels of mast cell-specific molecules contained by the lysates prepared with different numbers of PECs.
(A) PECs were stained with FITC-labeled anti-mouse c-kit plus PE-labeled anti-mouse IgE mAbs and analyzed with flow cytometry. Mast cells (i.e., c-kit+·IgE+ double positive cells) are shown in the upper-right quadrant. (B) Cell lysates were prepared with 1 x 106 (circle), 5 x 105 (square), and 2.5 x 105 (diamond) PECs, respectively, and one half of the total lysates was incubated with the substrate of β-hexosaminidase for a period of time as indicated. The extent of color changes by the enzyme reaction was measured with a 96 well microplate reader at 405 nm wavelength. Assays were conducted in duplicate. (C) Cell lysates were prepared as in (B) and the histamine concentrations in the lysates were measured with ELISA. (D) Mast cell-depleted PECs were stained with the mAbs as in (A). (E) Cell lysates were prepared with 5 x 105 crude (filled bar) and mast cell-depleted (open bar) PECs, respectively, and incubated with the β-hexosaminidase substrate for 30 min before termination of the reactions. (F) The concentrations of histamine in the cell lysates prepared as in (E) were measured. Please click here to view a larger version of this figure.
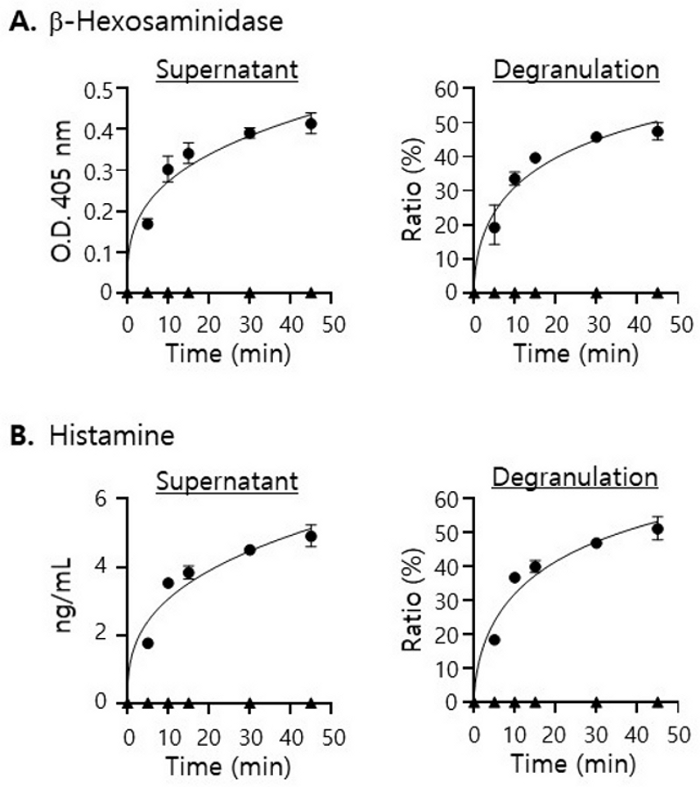
Figure 2: Measuring the amounts of mast cell-specific molecules released by mast cells during culture of PECs with DNP-BSA.
PECs (5 x 105) isolated from the mice injected with anti-DNP IgE (circle) or with just 1 x PBS (triangle) were cultured with DNP-BSA for a period of time as indicated. The levels of β-hexosaminidase (A, left) and histamine (B, left) detected in the supernatants at each time point were plotted. Extents of degranulation were calculated with the amounts of β-hexosaminidase (A, right) and histamine (B, right) released to the supernatants and remaining inside the cells (cell lysate), respectively. The following equation was used for calculating the extent of degranulation (%). Degranulation (%) = [X]sup /([X]sup + [X]lysate) X 100 Please click here to view a larger version of this figure.
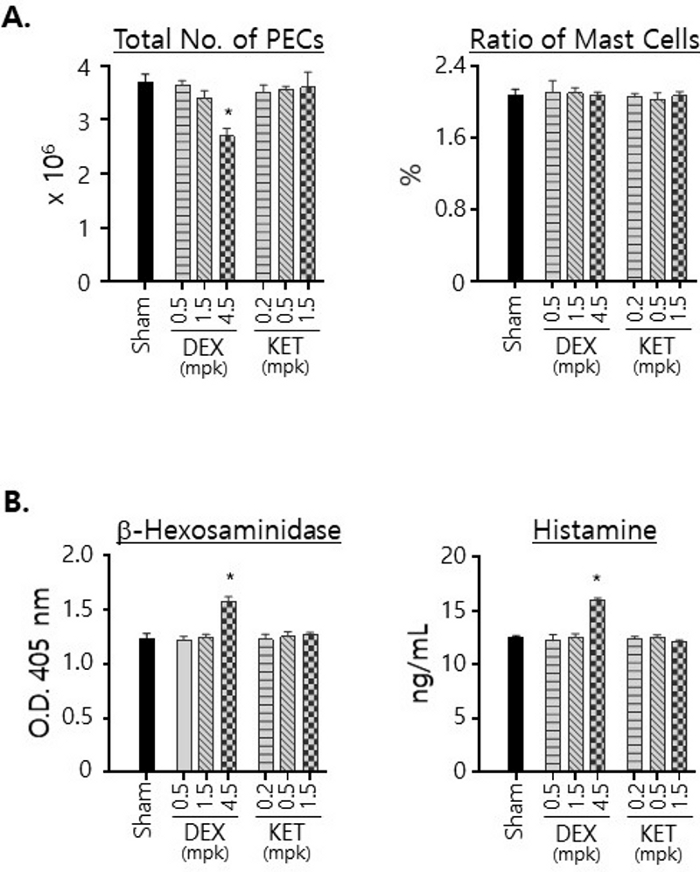
Figure 3: Effects of DEX and KET on the viability of mast cells in the peritoneal cavity and the levels of b-hexosaminidase and histamine expressions.
(A) PECs were isolated from the mice treated p.o. for 3 days with vehicle alone (sham) or with indicated doses of DEX or KET. Total cell numbers were counted with a hemocytometer and the average numbers were calculated and plotted along with standard deviations (left). PECs were stained with FITC-labeled anti-c-kit plus PE-labeled anti-IgE mAbs and analyzed with flow cytometry. The ratios of the double positive cells (i.e., mast cells) to total PECs were plotted (right). (B) PECs were isolated as in (A), and cell lysates were prepared with the same number (5 x 105) of PECs. The amounts of β-hexosaminidase (left) and histamine (right) in those lysates were measured and plotted. All experiments were performed with 3 mice per group. Statistical significances were calculated using one-way anova; * p < 0.05. Please click here to view a larger version of this figure.
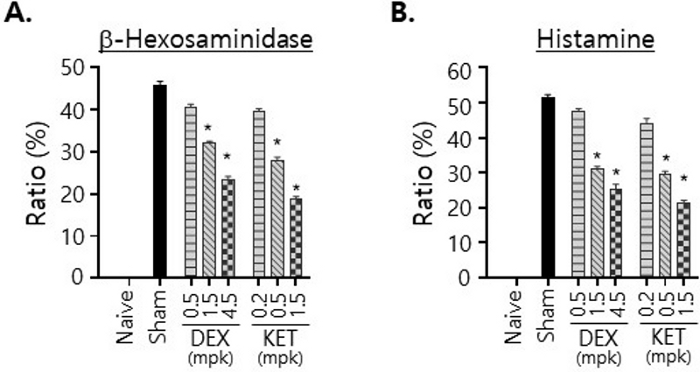
Figure 4: Ex vivo mast cell degranulation assay using crude PECs isolated from the mice treated with DEX or KET.
PECs were isolated from the mice treated p.o. for 3 days with vehicle alone (naïve and sham) or with indicated doses of DEX or KET and then injected i.v. with PBS (naïve) or anti-DNP-IgE. They were then incubated with DNP-BSA for 10 min, and the extents of degranulation were calculated based on the levels of β-hexosaminidase (A) and histamine (B), respectively, released to the culture supernatants. All experiments were performed with 3 mice per group. Statistical significances were calculated using one-way anova; * p < 0.05. Please click here to view a larger version of this figure.
| Buffer | Recipe | Comments/Description | |
| 0.1 M Na-citrate buffer | 45.6 mM Sodium Citrate dihydrate, 54.4 mM Citric Acid, pH 4.5 | Store at 2-8°C | |
| 10x RBC lysis buffer | 80 mg/mL NH4Cl, 8.4 mg/mL NaHCO3, 3.7 mg/mL EDTA (disodium) | Store at 2-8°C, do not exceed six months | |
| Cell lysis buffer | 0.1 % Triton X-100, 1 x phosphate-buffered saline (PBS) pH 7.4 | Store at 2-8°C | |
| ELISA stop solution | 100 mM glycine, pH 10.7 | Normal temperature storage | |
| FACS buffer | 1 x PBS, 5 % horse serum, 1 % BSA, 10 mM HEPES, 2 mM EDTA, pH 7.2 | Store at 2-8°C, do not exceed two months | |
| PBSBE buffer (MACS cell purification buffer) | 1x PBS, 0.5 % BSA, 2 mM EDTA, pH 7.2 | Store at 2-8°C, do not exceed two months | |
| Tyrode's A buffer | 10 mM HEPES, 130 mM NaCl, 5.6 mM glucose, 5 mM KCl, 1 mM MgCl2, 1.4 mM CaCl2, 1 % bovine serum albumin (BSA) pH 7.4 |
Store at 2-8°C, do not exceed two months | |
| Tyrode's B buffer | 137 mM NaCl, 5.6 mM glucose,12 mM NaHCO3,2.7 mM KCl, 0.3 mM NaH2PO4, 1 mM MgCl2, 1.8 mM CaCl2, 0.1 % gelatin,pH 7.4 | Store at 2-8°C, do not exceed two months | |
Table 1: Buffer compositions.
Supplemental Figure 1: Effects of DEX and KET on the anaphylactic reactions caused by passively transferred anti-DNP-IgE and DNP-BSA. Mice, treated (p.o.) daily for 3 days with the vehicle alone (naïve, sham) or with indicated doses of DEX or KET, were administered (i.v.) PBS (naive) or anti-DNP-IgE mAb. One day after the antibody injection, the mice were injected (i.v.) with PBS (naive) or with DNP-BSA. The body temperatures of the mice measured 30 min after DNP-BSA injection were plotted (A). One day after injection of DNP-BSA, the blood was drawn from the mice and the levels of MCPT-1 were measured with ELISA (B). The experiment was performed with 3 mice per group. Statistical significances were calculated using one-way ANOVA; * p < 0.05. Please click here to download this figure.
