Благодаря все более широкому применению методов секвенирования следующего поколения, пространственное разрешение в биологии развития может быть увеличено. Современные исследования направлены на вскрытие тканей вплоть до специализированных типов клеток, если не одноклеточного уровня1,,2,,3,,4. С этой целью за последние пятьдесят лет было разработано множество различных методов (см. рисунок 1A)5,6,6,7,,88,9,,10,,11,,12,,13,,14,,15.
Многие инструменты в науке о растениях были адаптацией методов, которые были впервые в исследованиях на животных. Это не относится к методу, который мы вводим в деталях здесь. В 2005 году, оснащенный сильным фоном в переводе белка, Лаборатория Бейли-Серрес приступила к разработке рибосомных белков для последующего очищения сродства16. Таким образом, они могли бы избежать трудоемких и трудоемких полисомного профилирования, которое основано на ультрацентрифугации с градиентом сахарозы и было использовано для оценки перевода рибосом с 1960-хгодов 17,18. Метод с тех пор был назван переводной рибосомы очистки сродства (TRAP)16. После успешных исследований транслатома в растениях, Heiman et al. адаптированы TRAP для животных19 и другие расширили свое применение дрожжей20, Drosophila21, Xenopus22 и зебрафиш23,24.
Хотя генетическая модификация модельной системы является необходимым условием для TRAP, которая ограничивает ее применение видов, поддавшихся генетической трансформации, можно одновременно использовать это возражение в целевых подмножествах клеток, которые представляют особый интерес и в противном случае чрезвычайно трудно изолировать от нетронутой ткани / органа25 (например, высоко разветвленные дендритные клетки в мозге мыши или грибковых hyphae инфицированных растительных тканей). В растениях все клетки удерживаются на месте через клеточные стенки, которые составляют основу гидростатического скелета26. Чтобы освободить клетку растений из этой матрицы, ученые либо физически вырезать клетку из окружающей ткани с помощью лазерного захвата микродиссекции (LCM)27 или выполняется ферментативное пищеварение клеточныхстенок 28. Среди последних клеток, так называемых протопластов, популяция, интересующая сяртово,констачированная и может быть отделена с помощью флуоресцентной сортировки клеток (FACS)7. LCM обычно требует, чтобы образец был исправлен и встроен в воск, что в конечном итоге ухудшает качество его РНК29. Методы на основе FACS дают высококачественную РНК, но сам процесс протоптертора вводит различия в экспрессии генов30, а ткани с модифицированными и толстыми вторичными клеточными стенками, как известно, трудно поддаются лечению. Кроме того, многие процессы развития в растениях, как предполагается, полагают, полагаются на механически передаваемые сигналы и, следовательно, целостность клеточной стенки имеет первостепенное значение31. Два метода, которые используют ярлык, чтобы обойти изоляцию клеток, работая на уровне ядер, являются флуоресценция активированной ядерной сортировки (FANS) и изоляции ядер помечены в конкретных типах клеток (INTACT). Как и в TRAP, они используют клеточные тип-специфические промоутеры для того чтобы маркировать ядра, которые затем получают обогащенными через сортировать или вытягивать вниз, соответственно8,,15. Основная проблема для всех этих подходов заключается в том, чтобы получить достаточное количество РНК-материала из подмножества клеток в ткани. Поскольку TRAP захватывает лишь часть клеточных РНК, сбор образцов является значительным узким местом. Поэтому для получения высококачественных данных из низких объемов ввода необходимы особенно чувствительные протоколы подготовки библиотек.
С момента своего создания, TRAP был либо использован в сочетании с ДНК microarrays или, как секвенирование расходы значительно снизились в последние годы, РНК-сек10,32,33. Множество вопросов исследования уже были выяснены, как рассмотрено в Sablok и др.34. Мы убеждены, что больше докладов будет следовать в ближайшие годы, как техника является очень универсальным при объединении различных промоутеров для целевых конкретных типов клеток. В конце концов, это будет сделано даже в индуцируемой образом, и может быть объединена с зондирования реакции растения на многие биотических и абиотических факторов стресса. Кроме того, там, где стабильные трансгенные линии отсутствуют, волосатые системы экспрессии корней также успешно используются для выполнения TRAP в томате и medicago35,36.
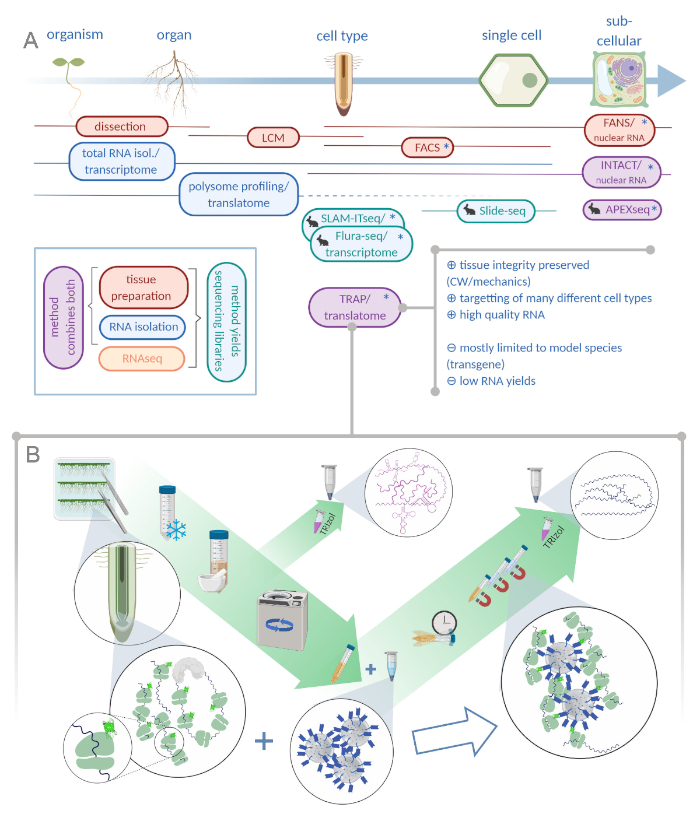
Рисунок 1: Перевод циризующей сродства рибосомы (TRAP) дополняет портфель анализа “омики”. О. Повышение уровня аналитической точности, вплоть до одноклеточного или даже субклеточного разрешения может быть достигнуто с помощью множества методов или их комбинаций. Схема дает обзор имеющихся в настоящее время инструментов в области растений и животных. Сбор тканей при клеточном разрешении может быть достигнут с помощью таких протоколов, как LCM или FACS, которые затем соединяются со стандартным транскриптомом или полисомным профилированием/анализом транслатома. TRAP и INTACT интегрируют захват тканей и изоляцию РНК, поскольку они основаны на эпитоп-пометке. Тем не менее, образцы INTACT только ядра клеток и представляет собой, следовательно, особый случай анализа транскриптома. Небольшой значок кролика отмечает недавно разработанные методы в области животных: В то время как SLAM-ITseq и Flura-seq полагаются на метаболическое тарзание зарождающихся РНК с модифицированными базами урацила в клетках, выражающих разрешительный фермент, Slide-seq использует стеклянный слайд с покрытием со штрих-кодами ДНК, которые предоставляют позиционную информацию в клеточном диапазоне. В APEX-seq используется подход к маркировке непосредственной маркировки для пробы РНК в конкретных субклеточных отсеках. Примечательно, что повышенное разрешение часто требует генерации трансгенного материала (звездочек), и эти методы, таким образом, в основном используются для типовых видов. TRAP особенно подходит для исследований растительной науки с участием клеточной стенки (CW) или механической сигнализации, а также клеточных видов, которые трудно выпустить из их матрицы CW. B. Детальные шаги влажной лаборатории процедуры TRAP: Рассада, выражающая GFP-тегами рибосомный белок в различных типах клеток (например, корневые эндодермисы), выращиваются на чашках Петри в течение семи дней, а корневой материал собирают путем замораживания оснастки. Общий образец контроля РНК собирается из гомогенизированного сырого экстракта, прежде чем гранулировать мусор с помощью центрифугации. Магнитные бусы анти-GFP добавляются в очищенный экстракт для выполнения иммунопротеберации. После инкубации и трех этапов стирки, полисомо-ассоциированной РНК (TRAP /полисома РНК) непосредственно получается через экстракцию фенола-хлороформа. LCM: лазерный захват микродиссекции, FACS/ FANS: флуоресценция активированной ячейки / ядерной сортировки, APEX-seq: метод, основанный на инженерии аскорбат перекистой, INTACT: изоляция ядер помечены в конкретных типах клеток, SLAM-ITseq: тиол (SH)-связанных алкилирования для метаболического секвенирования РНК в тканях, Flura-seq: фторурацил помечены РНК секвенирования (Создано с Biorender.com) Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
Целью данной статьи является предоставление подробного описания метода TRAP, выделение критических шагов и руководство по возможному методу подготовки библиотеки.
Общий эксперимент TRAP будет по существу состоять из следующих шагов (см. также Рисунок 1B):(1) Подготовка растительного материала, включая клонирование рибосом-теговая конструкция, трансгенное производство линии и отбор, выращивание и ссыпая семян, стерилизации и покрытия, а также стрессовое применение/обработка (необязательно) и сбор тканей; (2) иммуноочищение, включая гомогенизацию тканей и очистку сырого экстракта, мытье биса и иммуноочищение, и шаги мытья; (3) добыча РНК и оценка качества; и (4) подготовка библиотеки.
Корень Arabidopsis была модельсистемы для изучения развития растений с момента его введения в качестве модели завода37,38. Здесь применение TRAP демонстрируется в контексте развития бокового корня растений. В растениях, накопление всей корневой системы опирается на выполнение этой программы и поэтому очень важно для выживания организма39. В Arabidopsis, боковые корни происходят из ткани перицикл, который находится рядом с ксилем судов и, следовательно, называется ксилем полюс перицикл (XPP; см. Рисунок 2C)40. Некоторые клетки XPP, которые расположены глубоко внутри корня, приобретают идентичность ячейки основателя и, на местном гормональном триггере, начинают размножаться путем отеков и деления антиклинально41. Однако, из-за наличия жесткой матрицы стенки клетки, этот процесс оказывает механическое давление на окружающие ткани. В частности, поражена надлежащая эндодермис, так как она находится на пути боковой оси роста корня42,,43,,44. Действительно, вновь формирующихся primordium придется расти через надлежащую клетку эндодермис(Рисунок 2C2) в то время как коры и эпидермиз клетки просто отодвинуты в сторону для приморского,наконец,выйти45,46. Недавняя работа в нашей лаборатории показала, что эндодермис активно способствует распространению в перицикле. Целенаправленной блокировки эндодермальной гормональной сигнализации достаточно, чтобы ингибировать даже самое первое деление в клетках XPP47. Таким образом, перицикл эндодермис связи представляет собой очень ранний контрольно-пропускной пункт для бокового развития корней в Arabidopsis. Это, однако, не известно, как этот перекрестный разговор выполняется. Чтобы разгадать эту тайну, мы выбрали trap-seq подход к целевой XPP и эндодермальных клеток. Чтобы обогатить клетки в боковой корневой программе, мы имитировали гормональный триггер, экзогенно применяя аналог auxin (1-нафталенеацетической кислоты, NAA)48, который в то же время позволил временно решить начальную фазу бокового образования корня.
