Impulsionada pela crescente aplicação de técnicas de sequenciamento de última geração, a resolução espacial na biologia do desenvolvimento poderia ser aumentada. Estudos contemporâneos visam dissecar tecidos para tipos de células especializadas, se não o nível de célulaúnica11,2,,3,4. Para isso, uma infinidade de métodos diferentes foi concebida ao longo dos últimos cinquenta anos (ver Figura 1A)5,6,,7,,688,99,10,,11,,12,,13,,14,15.
Muitas ferramentas em ciência vegetal têm sido adaptações de técnicas que foram pioneiras na pesquisa animal. Este não é o caso para o método que estamos introduzindo em detalhes aqui. Em 2005, equipado com uma forte formação em tradução de proteínas, o Laboratório Bailey-Serres começou a projetar proteínas ribossômicas para posterior purificação de afinidade16. Assim, poderiam evitar o demorado e intensivo em perfil de polisério, que se baseia na ultracentrifugação com um gradiente de sacarose e foi utilizado para avaliar ribossomos traduzidos desde a década de 196017,18. Desde então, o método foi referido como purificação de afinidade de ribossomos traláricos (TRAP)16. Após estudos bem-sucedidos de translatome em plantas, Heiman et al. adaptaram TRAP para animais19 e outros estenderam sua aplicação para leveduras20, Drosophila21, Xenopus22 e zebrafish23,24.
Embora a modificação genética do sistema modelo seja um pré-requisito para trap, que limita sua aplicação a espécies passíveis de transformação genética, pode-se simultaneamente aproveitar essa objeção para atingir subconjuntos de células que são de interesse especial e de outra forma extremamente difíceis de isolar do tecido/órgão intacto25 (por exemplo, células dendríticas altamente ramificadas em um cérebro de camundongos ou hifas fúngicas em tecido vegetal infectado). Nas plantas, todas as células são mantidas no lugar através de paredes celulares que formam a base do esqueleto hidrostático26. Para libertar uma célula vegetal dessa matriz, os cientistas cortaram fisicamente a célula de seu tecido circundante através da microdissecção de captura a laser (LCM)27 ou realizaram digestão enzimática das paredes celulares28. Entre as últimas células, os chamados protoplastos, a população de interesse é fluorescentemente rotulada e pode ser separada por meio da triagem celular ativada por fluorescência (FACS)7. O LCM geralmente requer uma amostra para ser fixada e embutida em cera, o que acaba por deteriorar a qualidade do seu RNA29. Os métodos baseados em FACS produzem RNA de alta qualidade, mas o processo de protoplasting em si introduz diferenças na expressão genética30 e tecidos com paredes de células secundárias modificadas e grossas são notoriamente difíceis de tratar. Além disso, muitos processos de desenvolvimento nas plantas são assumidos para contar com sinais mecanicamente transmitidos e, portanto, a integridade da parede celular é de suma importância31. Dois métodos, que utilizam um atalho para contornar o isolamento celular operando no nível de núcleos, são a triagem nuclear ativada por fluorescência (FANS) e o isolamento de núcleos marcados em tipos de células específicas (INTACT). Como no TRAP, eles usam promotores específicos do tipo celular para marcar núcleos, que posteriormente são enriquecidos através da classificação ou puxar para baixo, respectivamente8,15. Um grande desafio para todas essas abordagens é obter material de RNA suficiente de subconjuntos de células em um tecido. Como o TRAP captura apenas uma fração dos RNAs celulares, a coleta de amostras é um gargalo considerável. Portanto, protocolos especialmente sensíveis de preparação de bibliotecas são necessários para produzir dados de alta qualidade a partir de baixas quantidades de entrada.
Desde a sua criação, o TRAP tem sido usado em combinação com micromatrizes de DNA ou, como os custos de sequenciamento caíram significativamente nos últimos anos, RNA-seq10,32,33. Uma infinidade de questões de pesquisa já foi elucidada como revisada em Sablok et al.34. Estamos convencidos de que mais relatórios serão seguidos nos próximos anos, pois a técnica é muito versátil ao combinar diferentes promotores para atingir tipos específicos de células. Eventualmente, isso será feito mesmo de forma indutível, e pode ser combinado com a sondagem da reação da planta a muitos fatores de estresse bióticos e abióticos. Além disso, quando linhas transgênicas estáveis não estão disponíveis, sistemas de expressão raiz peluda também têm sido usados com sucesso para realizar TRAP em tomate e medicago35,36.
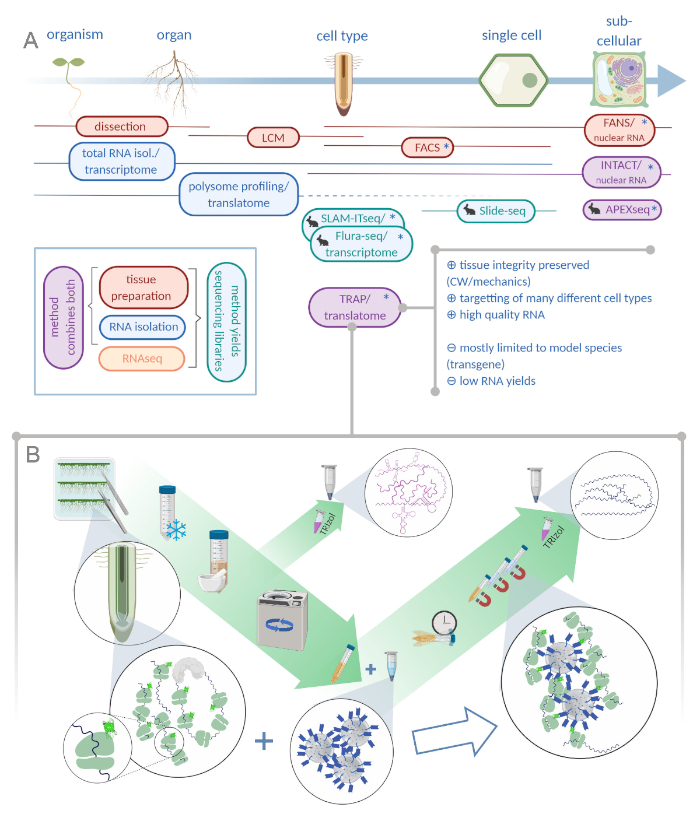
Figura 1: Traduzir purificação de afinidade de ribossomos (TRAP) complementa o portfólio de análise “ômica”. A. O aumento dos níveis de precisão analítica, até a resolução unicelular ou mesmo subcelular, pode ser alcançado por uma infinidade de métodos ou combinações. O esquema dá uma visão geral das ferramentas disponíveis atualmente no campo vegetal e animal. A coleta de tecidos na resolução celular pode ser alcançada por protocolos como LCM ou FACS, que são então acoplados à transcrição padrão ou análise de perfil/tradução polisome. TRAP e INTACT integram tanto a captura de tecido quanto o isolamento do RNA, pois são baseados na marcação de epítopos. No entanto, intacto amostra apenas núcleos celulares e constitui, portanto, um caso especial de análise de transcriptome. Um pequeno ícone de coelho marca métodos recém-desenvolvidos no campo animal: Enquanto SLAM-ITseq e Flura-seq dependem da segmentação metabólica de RNAs nascentes com bases uracil modificadas em células que expressam a enzima permissiva, Slide-seq faz uso de um slide de vidro revestido com códigos de barras de DNA que fornecem informações posicionais na faixa celular. Uma abordagem de rotulagem de proximidade é seguida em APEX-seq para amostrar RNAs em compartimentos subcelulares específicos. Notavelmente, o aumento da resolução muitas vezes requer a geração de material transgênico (asteriscos) e esses métodos são, portanto, predominantemente utilizados para espécies modelo. Trap é especialmente adequado para estudos de ciência vegetal envolvendo parede celular (CW) ou sinalização mecânica, bem como espécies celulares que são difíceis de liberar de sua matriz CW. B. Etapas detalhadas do laboratório molhado do procedimento TRAP: As mudas que expressam proteína ribossômica marcada por GFP em tipos de células distintas (por exemplo, endoderme raiz) são cultivadas em placas de Petri por sete dias e o material raiz colhido pelo congelamento instantâneo. Uma amostra total de controle de RNA é coletada do extrato bruto homogeneizado antes de pelleting os detritos via centrifugação. Contas anti-GFP magnéticas são adicionadas ao extrato limpo para realizar a imunoprecipitação. Após a incubação e três etapas de lavagem, o RNA associado ao pórnio (Trap/Polysome RNA) é obtido diretamente através da extração de fenol-clorofórmio. LCM: microdissecção de captura a laser, FACS/FANS: célula ativada por fluorescência/classificação nuclear, APEX-seq: método baseado na engenharia de ascorbate peroxidase, INTACT: isolamento de núcleos marcados em tipos de células específicas, SLAM-ITseq: alquilação vinculada ao thiol(SH) para o sequenciamento metabólico de RNA no tecido, Flura-seq: sequenciamento de RNA com rótulo fluorouracil (Criado com Biorender.com) Clique aqui para ver uma versão maior desta figura.
O objetivo deste artigo é fornecer uma descrição detalhada do método TRAP, destacar passos críticos e fornecer orientação para um possível método de preparação da biblioteca.
Um experimento trap genérico consistirá essencialmente nas seguintes etapas (ver também Figura 1B): (1) Preparação de material vegetal incluindo clonagem de construção de marcação de ribossomos, produção e seleção de linhas transgênicas, crescimento e acúmulo de sementes, esterilização e revestimento, e aplicação/tratamento de estresse (opcional) e colheita de tecidos; (2) imunopurificação, incluindo homogeneização tecidual e limpeza do extrato bruto, lavagem de abelhas e imunopurificação, e etapas de lavagem; (3) Extração de RNA e avaliação da qualidade; e (4) preparação da biblioteca.
A raiz arabidopsis tem sido um sistema modelo para estudar o desenvolvimento de plantas desde sua introdução como uma planta modelo37,38. Aqui, a aplicação do TRAP é apresentada no contexto do desenvolvimento da raiz lateral vegetal. Nas plantas, o acúmulo de todo o sistema radicular depende da execução deste programa e, portanto, é muito importante para a sobrevivência do organismo39. Em Arabidopsis,as raízes laterais originam-se do tecido periciclo que reside ao lado dos vasos xilem e, portanto, é denominado periciclo do pólo xilem (XPP; ver Figura 2C)40. Algumas células XPP, que estão localizadas no interior da raiz, adquirem uma identidade celular fundadora e, sobre um gatilho hormonal local, começam a se proliferar por inchaço e divisão anticlinalmente41. No entanto, devido à presença de uma matriz de parede celular rígida, este processo exerce estresse mecânico nos tecidos circundantes. Em particular, a endoderme sobreposta é afetada, pois está no caminho do eixo de crescimento da raiz lateral42,43,44. De fato, o primordium recém-formado terá que crescer através da célula endoderme sobreposta (Figura 2C2), enquanto as células córtex e epiderme são apenas deixadas de lado para que o primordium finalmente emerja45,46. Trabalhos recentes em nosso laboratório mostraram que a endoderme está contribuindo ativamente para acomodar a proliferação no periciclo. O bloqueio direcionado da sinalização hormonal endodérmica é suficiente para inibir até mesmo a primeira divisão nas células XPP47. Assim, a comunicação periciclo-endoderme constitui um ponto de verificação muito cedo para o desenvolvimento da raiz lateral na Arabidopsis. No entanto, não se sabe como esse crosstalk é realizado. Para desvendar esse mistério, escolhemos a abordagem TRAP-seq para atingir células XPP e endodérmicas. Para enriquecer para as células no programa raiz lateral, imitamos o gatilho hormonal aplicando exogenously um análogo auxina (1-ácido naftalinacático, NAA)48, que ao mesmo tempo permitiu resolver temporalmente a fase inicial da formação da raiz lateral.
