L'interaction entre les molécules est la base de la nature. Par conséquent, les scientifiques dans de nombreux domaines de la recherche fondamentale et appliquée tentent de comprendre les principes fondamentaux des interactions moléculaires de différents types. MicroScale thermophorèse (MST) permet aux scientifiques d'effectuer la caractérisation précise rapide, rentable et de qualité contrôlée des interactions moléculaires en solution, avec un libre choix de tampons. Il y a déjà plus de 1000 publications en utilisant MST, à partir de 2016 seulement, décrivant les différents types d'analyses, y compris des projections de la bibliothèque, les validations d'événements, des essais de compétition, et les expériences de liaison avec plusieurs partenaires de liaison 1-8. En général, le MST permet l'étude des paramètres de liaison classiques, tels que l'affinité de liaison (pM mM), stoechiométrie, et de la thermodynamique, de tout type d'interaction moléculaire. Un grand avantage de MST est la possibilité d'étudier des événements de liaison indépendants de la taille des partenaires d'interaction. Même chalinteractions lenging entre les petites aptamères d'acides nucléiques (15-30 nt) et des cibles telles que des petites molécules, des médicaments, des antibiotiques, ou métabolites peuvent être quantifiés.
Les technologies actuelles state-of-the-art pour caractériser les interactions aptamère-cibles sont soit en laboratoire intense et très complexe ou ne parviennent pas à quantifier les interactions aptamère-petite molécule 9,10. Surface Plasmon Resonance (SPR) à base de dosages 11,12 et approches colorimétriques vraiment sans étiquette, comme Calorimétrie titration isotherme (ITC) 13-15, élution isocratique 16, l' équilibre fi ltration 17,18, en ligne de sondage 19, gel – passer des tests, arrêt – fl ux fl uorescence spectroscopie 20,21, anisotropie de fluorescence (FA) 22,23, une seule molécule fl uorescence imagerie 24,25, et Bio-couche interférométrie (BLI) 26 sont également soit imprécis ou incompatible avec la molécule d'aptamères-petite interactions. Autres principal enjeux de ces méthodes sont une faible sensibilité, une consommation élevée de l'échantillon, l'immobilisation, les limitations de transport de masse sur les surfaces, et / ou des restrictions de mémoire tampon. Seules quelques-unes de ces technologies offrent des contrôles intégrés pour l'agrégation et d'adsorption des effets.
MST représente un outil puissant pour les scientifiques de surmonter cette limitation pour étudier les interactions entre les aptamères et les petites molécules 27-29, ainsi que d' autres cibles telles que les protéines 30-33. La technologie repose sur le mouvement de molécules à travers des gradients de température. Ce mouvement dirigé, appelé "thermophorèse," dépend de la taille, la charge, et la coquille d'hydratation de la molécule 34,35. La liaison d'un ligand à la molécule directement altérer au moins l'un de ces paramètres, ce qui entraîne une mobilité thermophorétique changé. Ligands avec de petites tailles peuvent ne pas avoir un impact considérable en termes de changement de taille de la non liée à l'état lié, mais ils peuvent avoir dr effets Amatic sur la coque et / ou la charge hydratation. Les changements dans le mouvement thermophorétique des molécules après interactions avec le partenaire de liaison permet la quantification des paramètres de liaison de base 2,7,34,36,37.
Comme cela est représenté sur la figure 1A, le dispositif MST est constitué d'un laser infrarouge focalisé sur l'échantillon à l' intérieur des tubes capillaires en verre optique en utilisant les mêmes que pour la détection de la fluorescence. Le mouvement thermophorétique de protéines via le fl uorescence intrinsèque de tryptophanes 6 ou d'un partenaire d' interaction 3,8 marqué par fluorescence peut être surveillée tandis que le laser établit un gradient de température (AT de 2-6 ° C). La différence de température dans l' espace, AT, conduit à l'épuisement ou l' accumulation de molécules dans le domaine de température élevée, qui peut être quantifiée par la Soret coef fi cient (S T):
g "/>
c représente la concentration à chaud dans la région chauffée et froid c est la concentration initiale dans la zone froide.
Comme le montre la figure 1B, un exemple typique de résultats expérimentaux MST dans un profil de mouvement MST (trace de temps), composé de différentes phases, qui peuvent être séparés par leurs échelles de temps respectives. La fluorescence initiale est mesurée au cours des 5 premières secondes en absence de gradient de température pour définir la fluorescence de départ précis et pour vérifier photoblanchiment ou photoenhancement. Le saut de température (T-Jump) représente la phase dans laquelle les changements de fluorescence avant le mouvement thermophorétique. Cette diminution initiale de la fluorescence dépend des changements de chaleur dépendant de fl uorophore rendement quantique. La phase de thermophorèse suit, dans laquelle les baisses de fluorescence (ou augmente) en raison du mouvement thermophorétique des molécules jusqu'à ce que la distribution à l'état d'équilibre est atteint.Le tjump inverse et rétrodiffusion concomitante de molécules fluorescents peuvent être observées comme indiqué dans la figure 1B après le laser est éteint. Pour accéder aux paramètres de liaison de base, différents rapports molaires des partenaires d'interaction sont analysées et comparées. En règle générale, 16 rapports différents sont étudiés dans une expérience MST, tandis que la molécule visible optique est maintenue constante et est alimentée avec une quantité croissante du ligand non marqué. L'interaction entre les deux partenaires de liaison induit des changements dans la thermophorèse, et donc dans la fluorescence normalisée, F norme, qui est calculé comme suit:
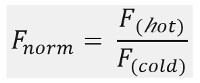
F chaud et froid F représentent en moyenne fl intensités uorescence au dé fi nies points de temps des traces du MST. Les affinités de liaison (K d ou CE 50 valeurs) peuvent être calculées par curve montage (figure 1C).
Dans l'ensemble, le MST est un outil puissant pour étudier les interactions moléculaires de toute nature. Ce manuscrit propose un protocole pour caractériser l'interaction difficile entre la petite triphosphate molécule d'adénosine (ATP; 0,5 kDa) et le 25-nt court ADNss aptamère DH25.42 (7,9 kDa). Au cours du manuscrit, le site de liaison de l'aptamère sur la molécule d'ATP est mappée vers le bas pour le groupe adénine de l'ATP.
