To form the root surface microstructure replication, a root must be chosen for molding. We grow tomato plants in soil, making the use of the natural root from the root system extremely challenging. Removal of soil from the root system can be difficult and additionally, the root system roots are fragile and can break upon molding attempts. We therefore suggest to first use more rigid roots, to establish the protocol in the lab. The formation of such roots is described in Figure 1A. The plant root system is removed after the plant was grown for 3 weeks and the rootless plant is placed in water for about a week until adventitious roots emerge from the stem. Those roots can be used for replication during the protocol establishment. Once the protocol has been well established, a more realistic root surface structure is desired. Here we suggest avoiding roots grown in soil as the full removal of soil in extremely challenging. Instead we suggest the use of germinating roots, supplying valuable information on the root surface microstructure of a genetically specific plant. The growth of such roots is described in Figure 1B. The seeds are placed on a wet filter paper and incubated at 25 °C. After approximately 5 days, during which the filter paper is kept moist, the germinated roots are long enough for replication. Those roots are more fragile than the previously suggested roots and require more delicate care.
The production of the root surface microstructure replica is a two-step process. In the first step the natural root is being molded into a polyurethane based mold (the negative replica). The advantage of this step is that all materials for the polyurethane mold are being prepared and the root is placed on top of the prepared solution at the very end for a 10 min exposure to UV. As a result, the biological tissue is not exposed to harsh conditions for too long and can be gently handled at the end of the process. If all protocol steps are followed, a good negative replica is generated. This replica will show the cell structure of the root surface as well as holes representing the location of the root hairs (Figure 2A). If some critical steps in the protocol are not being followed, the procedure will fail. One such step is the placement of the root on the polyurethane solution prior to curing. The root must be placed very gently to avoid the submergence of it in the polyurethane solution. Such submergence, of any part of the root, will cause the entrapment of the root in the hard polymer with no ability to remove it. If such an event occurs, the root will remain within the negative replica after it is cured (Figure 2B). Another crucial step is regarding the curing time by UV light. The recommended curing time is 8‒10 min. Going past 10 min will result in an extremely hard polyurethane mold, making it impossible to remove the root without breaking it within the polyurethane mold. The breakage of the root can sometimes be visible to the naked eye, e.g., when a large piece is broken (Figure 2C, top, marked with purple arrows). However, sometimes small root pieces are left in the material which are difficult to spot by the naked eye and a microscope has to be used (Figure 2C, bottom, marked with purple arrows). We recommend carefully examining the polyurethane negative replica with a microscope prior to the continuation of the protocol to make sure no residual root is present.
Once the polyurethane negative replica is prepared; many materials can be used for the preparation of the positive replica. The preparation of the positive replica, using the polyurethane negative replica as a mold, is straight forward and depends completely on the quality of the polyurethane negative replica. To generate the positive replica we have used both PDMS—as it is well known in the field of soft lithography (Figure 3A)—and ethyl cellulose as a material that better mimics the properties of the root surface which is mostly composed of cellulose (Figure 3B). The SEM image of the PDMS replica shows the root hairs very clearly. The hairs are in the elongation zone, where they begin to emerge. Hence, the length of root hairs varies along the root surface as they become longer, much like in the natural root (Figure 3A). Ethyl cellulose generates harder and less flexible film than PDMS. Hence, the removal of it from the negative mold requires more care. However, some hairs and the surface microstructure are visible under the light microscope (Figure 3B). We used those two materials to generate the positive replica, however, any material that can form a film will be a good candidate for the positive replica, using the polyurethane negative replica.

Figure 1: Tomato plant roots for replication. (A) A tomato (M82) plant is grown at 25 °C with 9 h of light and 15 h darkness. After 3 weeks, the plant is removed from the soil and the root system is cut off. The rootless plant is put in water until adventitious roots emerge from the stem after about a week. These roots do not show the exact structure as the roots from the root system, but they represent a good model. Those roots are less fragile than the root system roots and hence are preferred to work with when establishing the technique in the lab. (B) Tomato (M82) seeds are put on a wet filter paper in a Petri dish and incubated at 25 °C. The paper is hydrated every day and the seeds are germinating. The roots are growing and after approximately 5 days are long enough to be used for replication. These roots are gentler and should be used once the method is well established. Please click here to view a larger version of this figure.

Figure 2: Microscopy images of polyurethane negative replica. (A) SEM image of polyurethane negative replica made according to a protocol following all the steps. Cell structure is clearly visible. Yellow arrows point at holes formed by the hairs in the root. (B) Light microscopy images of polyurethane negative replica with a root inside of it as it was fully covered with the solution and the removal of it was impossible. The polyurethane negative was cured with the root inside. The root is visible by eye and using light microscopy. It is impossible to remove this root from the cured replica. (C) Light microscopy images of polyurethane negative replica that was kept under UV light for too long. As a result, root could not be fully removed from the polymer with either large particles visible by eye (top image, marked with purple arrows) or small fractions visible only by microscope (lower image, marked with purple arrows). Please click here to view a larger version of this figure.
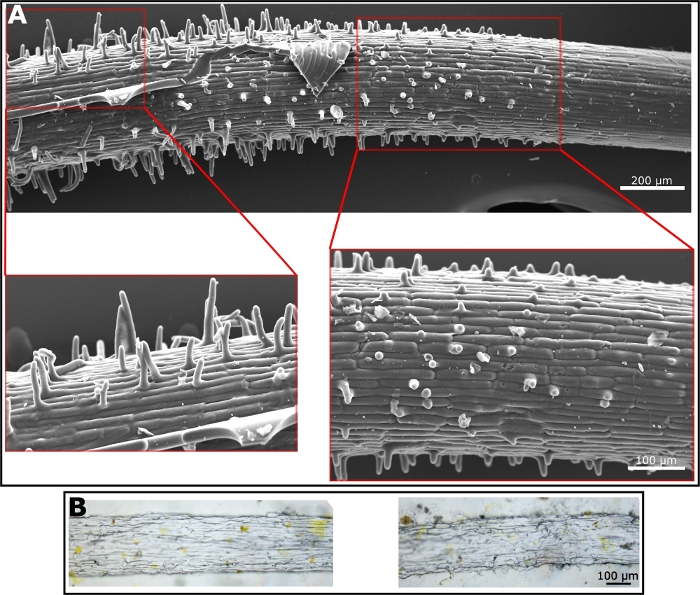
Figure 3: Microscope images of positive replica. (A) SEM micrograph of a positive replica made from PDMS. Enlargement shows root hairs. (B) Light microscopy images of a positive replica made from ethyl cellulose. Hairs are shown in the images on the right while surface texture is visible in the image on the left. Please click here to view a larger version of this figure.
