Figure 1 shows plates of PAO1 and PAO581 with or without in-frame deletion in the pyrD gene (a gene in the pyrimidine biosynthesis pathway) that results in SCV6. The PAO1 SCV mutant was restored to normal growth in response to uracil supplementation (Figure 1A,B). Furthermore, the PAO581ΔpyrDSCV mutant was returned to mucoidy with the same uracil treatment, because the parent strain PAO581 has an additional mucA25 mutation (Figure 1C,D). The results for the uronic acid carbazole assay are shown in Figure 26. The data represents samples of PAO581 and PAO581 with mutations in genes regulating pyrimidine de novo biosynthesis grown on PIA and PIA supplemented with 0.1 mM of uracil (Figure 2A). The data shows that the presence of uracil in the media results in the conversion of the mutant strain back to mucoidy (as seen by the increase/restored alginate production) (Figure 2B). The results for the anti-alginate monoclonal antibody based ELISA are represented in Figure 3. The data shows PAO1, PAO1 with an in-frame deletion of the algD gene encoding the key alginate biosynthetic enzyme GDP-mannose dehydrogenase, and PAO1 carrying an expression plasmid pHERD20T with the main alginate-specific sigma factor algU when grown on PIA plates with arabinose for the induction the pBAD promoter in pHERD20T. This data shows the non-mucoid levels of alginate measured for PAO1, and PAO1ΔalgD versus the mucoid levels of alginate measured for PAO1+pHERD20T-algU. Figure 4 compares the two methods of alginate measurements together. The results were not statistically significant when compared to each other using a two-way ANOVA with p < 0.01. Figure 5A compares the cross reactivity of the anti-alginate mAb against other polysaccharides including amylopectin, amylose, collagen, and glycogen. Figure 5B shows the comparison in specificity and sensitivity of the uronic acid carbazole assay to the newly developed anti-alginate monoclonal antibody-based ELISA with the control utilizing the highly purified seaweed alginate (Table of Materials). Figure 6 shows direct testing of the ELISA on patient sputum samples that were positive for mucoid P. aeruginosa and patients that did not contain mucoid P. aeruginosa.
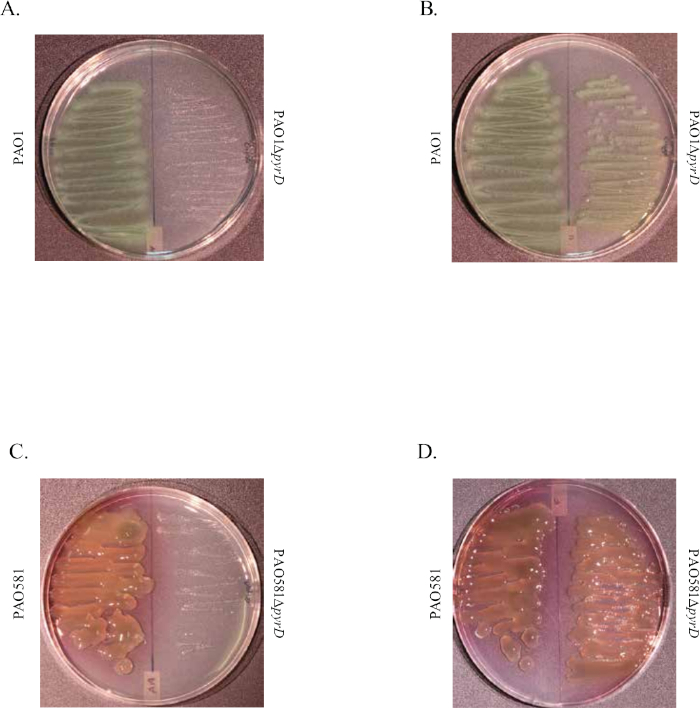
Figure 1: Representative images of the de novo pyrimidine biosynthesis mutations that result in SCV phenotype in PAO1 and PAO581. Image shows PAO1 (left) and PAO1ΔpyrD (right) on PIA plates (A) and PIA plates supplemented with uracil (B) grown at 37 °C for 48 h. Image shows PAO581 (left) and PAO581ΔpyrD (right) on PIA plates (C) and PIA plates supplemented with uracil (D) grown at 37 °C for 48 h. This figure has been modified from work done by Al Ahmar et al.6. Please click here to view a larger version of this figure.
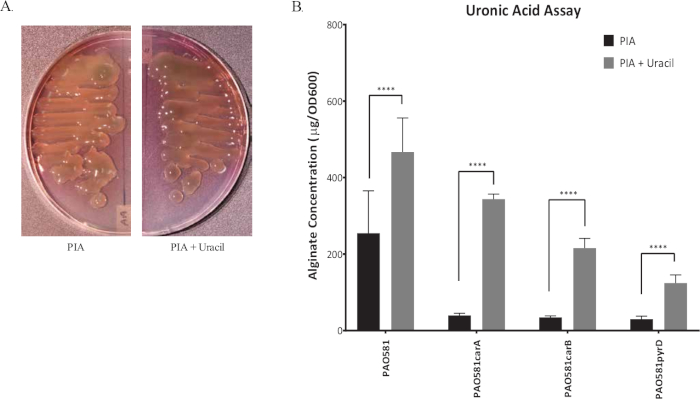
Figure 2: Representative graph of uronic acid carbazole assay. (A) The image of mucoid P. aeruginosa strain PAO581(PAO1mucA25) grown at 37 °C for 24 h on PIA plates (left) and PIA plates with uracil (right). (B) Alginate production for PAO581, PAO581carA, PAO581carB, and PAO581pyrD when grown on PIA plates with and without uracil at 37 °C for 24 h. Alginate was collected and measured using the standard carbazole assay. Values shown are mean alginate ± standard deviation of triplicate reads. (**** = p < 0.0001). This figure has been modified from work done by Al Ahmar et al.6. Please click here to view a larger version of this figure.
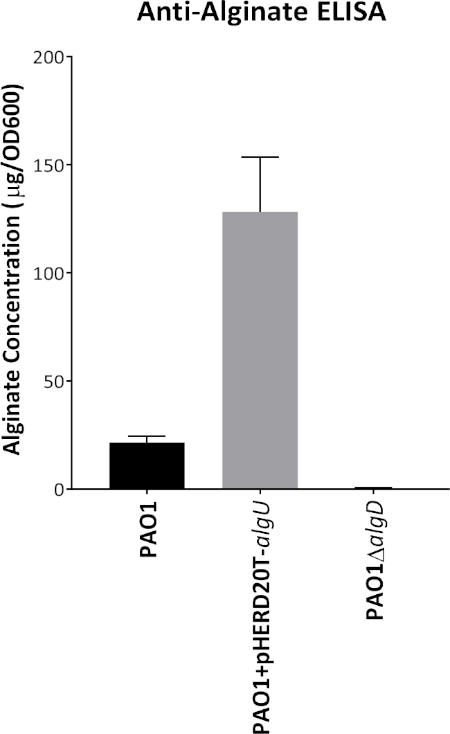
Figure 3: Representative graph of anti-alginate mAb-based ELISA. Alginate production for PAO1, PAO1ΔalgD and PAO1 carrying the expression vector pHERD20T-algU grown on PIA with 0.1% arabinose at 37 °C for 24 h. Alginate was collected and measured using the anti-alginate ELISA with the mouse anti-alginate monoclonal antibody. Values shown are mean alginate ± standard deviation of triplicate reads. ****p < 0.0001. Please click here to view a larger version of this figure.
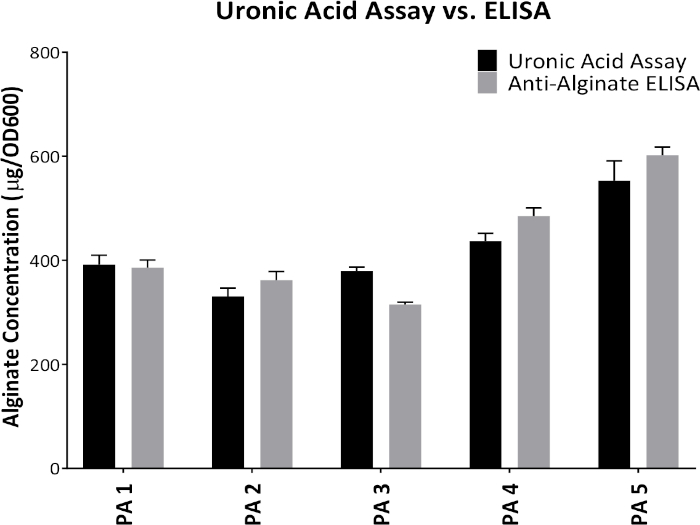
Figure 4: Comparison between the results obtained from uronic acid assay and anti-alginate mAb-based ELISA. Alginate production from five different mucoid P. aeruginosa proprietary strains grown on PIA plates at 37 °C for 24 h. Alginate was collected and measured by the uronic acid assay and anti-alginate ELISA. Values shown are mean alginate ± standard deviation of triplicate reads. Please click here to view a larger version of this figure.
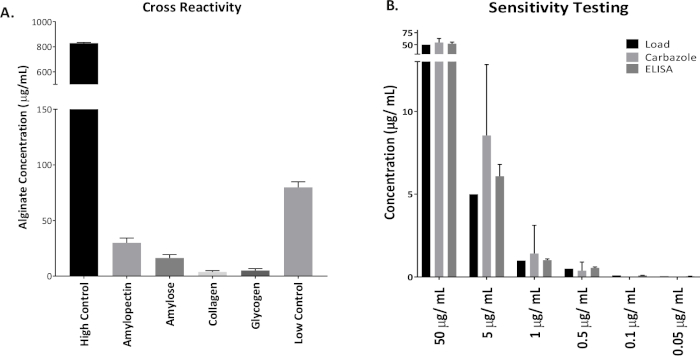
Figure 5: Specificity and sensitivity of the anti-alginate mAb based ELISA in comparison to the uronic acid carbazole assay. (A) ELISA was run with high (800 µg/mL) and low (100 µg/mL) internal assay controls of the seaweed alginate. This alginate was also used as a standard for the ELISA. Other polysaccharides tested that may cross react with anti-alginate mAb were amylopectin, amylose, collagen, and glycogen (500 µg/mL each). (B) Uronic acid carbazole assay and ELISA were run using the same range of standard concentrations with seaweed alginate: 50 µg/mL, 5 µg/mL, 1 µg/mL, 0.5 µg/mL, 0.1 µg/mL, and 0.05 µg/mL. Values shown are mean alginate ± standard deviation of triplicate reads. Please click here to view a larger version of this figure.
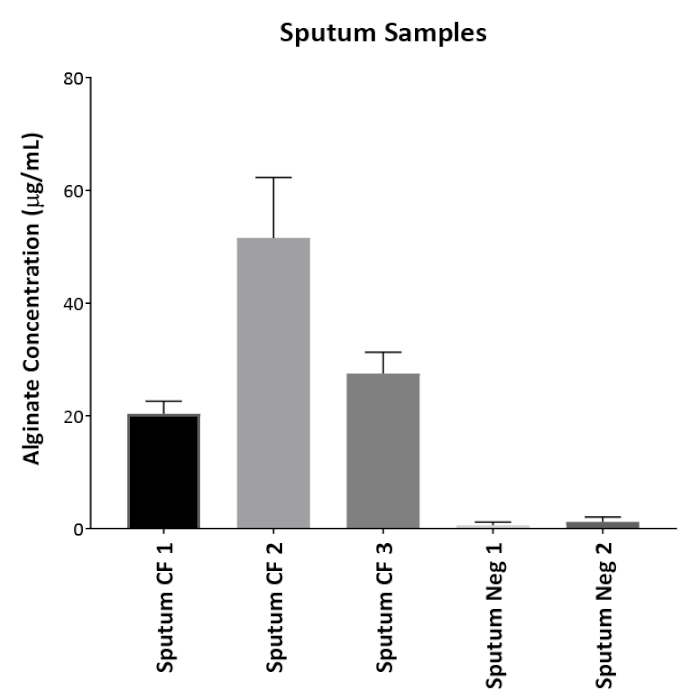
Figure 6: Direct patient sample testing. Anti-alginate ELISA was tested on patients' sputum samples without prior growth on plates. Three CF sputum samples that had growth of mucoid P. aeruginosa were used as well as two patient sputum samples that contained either non-mucoid P. aeruginosa (Neg 1) or no P. aeruginosa growth (Neg 2). Values shown are mean alginate ± standard deviation of triplicate reads. Please click here to view a larger version of this figure.
