Dynamic modification of 5mC in DNA by TET-family dioxygenases plays important roles in epigenetic transcriptional regulations. TET2 dioxygenase is frequently mutated in diverse hematopoietic malignancies12. To investigate the role of the TET2 enzyme in normal development and disease, we have cloned its minimal catalytically active domain without any affinity tag into the pDEST14 vector22. The untagged TET2 dioxygenase was produced at ∼5% of the total soluble protein by SDS-PAGE analysis in bacterial E. coli BL21 (DE3) cells. Since the catalytic domain of TET2 has a relatively high isoelectric point (∼7.49), compared with most indigenous E. coli proteins24,25,26, an efficient purification process utilizing a cation exchange chromatography was developed. This purification yielded >90% pure TET2 enzyme in a single step (Figure 1).
In order to separate and quantify different deoxycytidines derivatives and other four natural DNA bases following the TET2 enzymatic reaction, a sensitive LC-MS/MS-based assay was optimized. The liquid chromatography used a reversed-phase C18 columns. Standard curves were drawn using serial dilutions of a mixture containing all nucleosides (Figure 2). The gradient used for liquid chromatography, described in the experimental procedure, was able to resolve all eight nucleosides (Figure 3). The LC retention times (tr) for all eight nucleosides are described in Table 3. We further optimized the MS detection of each parent ion nucleoside (Q1), the most intense product ion (Q3) by determining their declustering potential (DP), entrance potential (EP), collision cell entrance potential (CEP), collision energy (CE), limit of detection (LOD), and lower limit of quantification (LLOQ) (Table 3). Finally, an LC-MS/MS method was developed that can separate and quantify the four normal DNA bases (A, T, G, and C), as well as the four modified cytosine bases (5-methyl, 5-hydroxymethyl, 5-formyl, and 5-carboxyl) (Figure 3).
The activity of untagged TET2 dioxygenase was determined using a 25-mer dsDNA containing one 5mC in a CpG island in each DNA strand (Table 2). After TET2 enzymatic reactions, DNA oligonucleotides were purified and converted into nucleosides. Then these nucleosides were subjected to LC-MS/MS assay. In the reactions without the TET2 enzyme (negative control), only dA, dT, dG, dC, and 5mdC peaks were observed. However, in the positive control reaction, which contained the TET2 dioxygenase, two new peaks corresponding to d5hmC and d5fC were observed. We were not able to detect the formation of d5caC nucleoside possibly due to its poor detection levels (Figure 2). These results demonstrate that the untagged TET2 dioxygenase purified in this procedure is catalytically active and can be used to characterize the wt-TET2 enzyme and its clinical mutants.
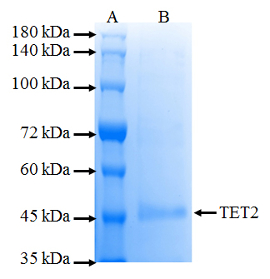
Figure 1. SDS-PAGE analysis of purified TET2 dioxygenase from E. coli BL21 (DE3) cells. Lane A indicates marker while lane B indicates TET2 protein purified using SP sepharose ion exchange resin. The total size of the untagged TET2 dioxygenase is ∼54 kDa as indicated by the arrow. Please click here to view a larger version of this figure.
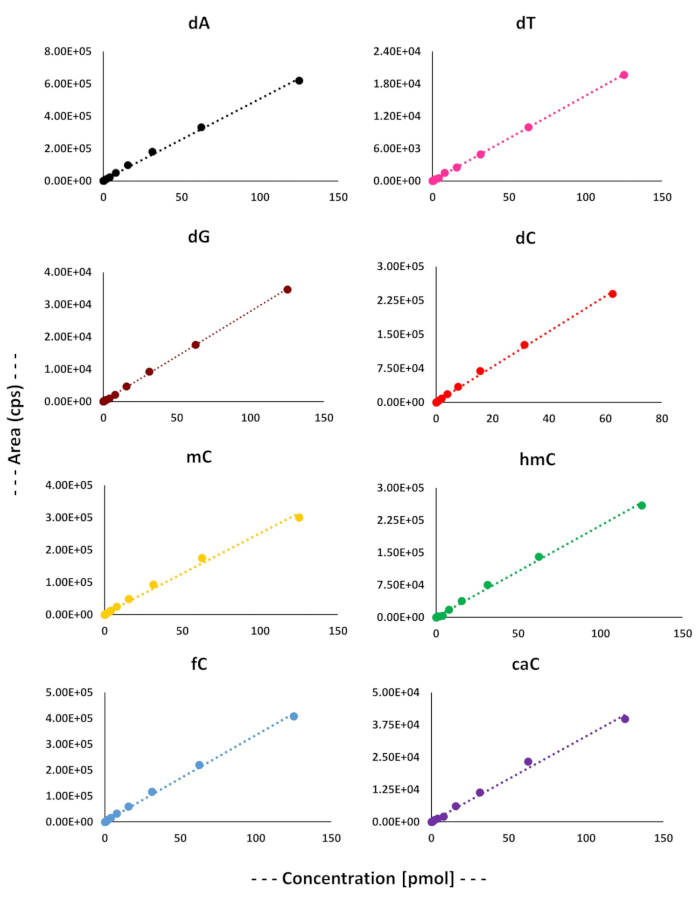
Figure 2. Standard curves was draw for four natural DNA nucleosides and different cytosine derivatives, which were then used for their quantification. Please click here to view a larger version of this figure.
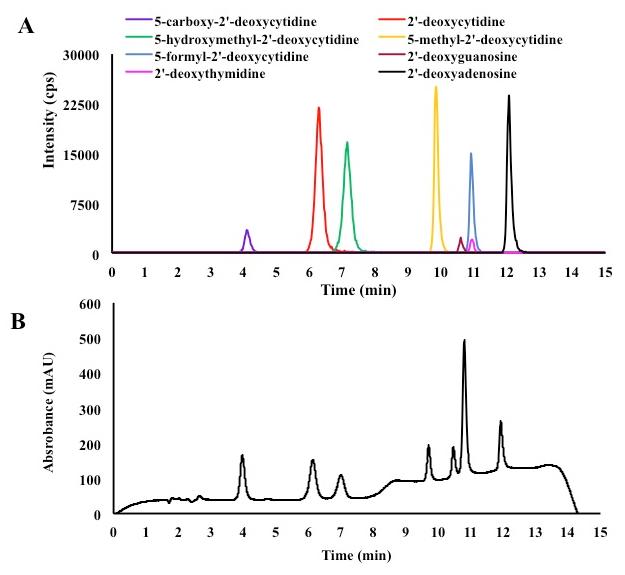
Figure 3. Liquid chromatography (bottom) and MS/MS (above) method used to separate and characterize four natural DNA nucleosides and different cytosine derivatives.
| Primer Name | Primer Sequence | |||
| TET2 forward primer | 5'-GGGGACAAGTTTGTACAAAAAAGCAGGCTTCGAAGGAGATAGAACCATGTCTGTTCTCAATAATTTTATAG-3' | |||
| TET2 Reverse Primer | 5'-GGGGACCACTTTGTACAAGAAAGCTGGGTCTCAGCCATACTTTTCACAC-3' | |||
Table 1: Sequence of DNA oligonucleotide primers used for PCR amplification of the catalytic domain of untagged human TET2 dioxygenase.
| Primer Name | Primer Sequence |
| Sense Strand | 5'-AGCCCGCGCCG/iMe-dC/GCCGGTCGAGCGG-3' |
| Antisense Strand | 5'-CCGCTCGACCGGCG/iMe-dC/GGCGCGGGCT-3' |
Table 2: Sequence of the sense and anti-sense 25-mer dsDNA oligonucleotide used as a TET2 substrate for in vitro oxidation reactions.
| Nucleosides | Q1 | Q3 | tr (min) | DP (V) | EP (V) | CEP (V) | CE (V) | LOD (pmol) | LLOQ | R2 | |
| 2'-deoxyadenosine | 252.2 | 136.1 | 12.07 | 41 | 9 | 14 | 17 | 0.06 | 0.198 | 0.997 | |
| 2'-deoxythymidine | 243.2 | 117.1 | 10.95 | 16 | 8 | 14 | 15 | 1.8 | 5.94 | 0.999 | |
| 2'-deoxyguanosine | 268.2 | 152.1 | 10.6 | 21 | 7 | 14 | 37 | 7.8 | 25.74 | 0.999 | |
| 2'-deoxycytidine | 228.1 | 112.1 | 6.29 | 21 | 7 | 14 | 15 | 0.1 | 0.33 | 0.998 | |
| 5-methyl-2'-deoxycytidine | 242.2 | 126.1 | 9.85 | 31 | 6.5 | 24 | 13 | 0.03 | 0.1 | 0.998 | |
| 5-hydroxymethyl-2'- deoxycytidine | 258.2 | 142.1 | 7.15 | 16 | 6 | 14 | 13 | 0.6 | 1.98 | 0.993 | |
| 5-formyl-2'-deoxycytidine | 256.2 | 140.1 | 10.92 | 11 | 6 | 14 | 15 | 0.2 | 0.66 | 0.998 | |
| 5-carboxy-2'-deoxycytidine | 272.2 | 156.1 | 4.1 | 6 | 7 | 94 | 23 | 3.9 | 12.87 | 0.993 | |
Table 3: Optimized LC-MS/MS parameters of four natural DNA nucleosides and different cytosine derivatives under positive ion mode. For each parent ion nucleoside (Q1), the most intense product ion (Q3) was detected.
