Het bepalen van de structuur van een biomolecuul is een belangrijke voorwaarde voor het begrijpen van zijn functie. Twee goed gevestigde methoden voor structuurbepaling zijn cryo-elektronenmicroscopie en X-ray kristallografie 1, 2. Vandaag, beide methoden bieden een hoge-resolutie structurele informatie met een resolutie tot aan de Angstrom niveau. Deze twee methoden zijn uitgebreid gebruikt om de structuur van grote biomoleculen zoals helderen eiwitcomplexen. Hoewel de bestaande werkwijzen continu gedurende de laatste jaren verbeterd, de complexiteit van biologische structuren vormt nog steeds een belangrijke uitdaging voor structurele biologie, in het bijzonder bij grote, dynamische en kortlevende complexen onderzocht 3.
Om de dynamiek van macromoleculaire complexen en de structuur-functie relatie name bestuderen single-molecule methodologieën provided nuttige informatie 4. Verschillende nieuwe strategieën ontwikkeld verschaffen van een orthogonale aanpak op het verwerven van structurele en dynamische informatie. Voorbeelden zijn hoge snelheid AFM 5, mechanische manipulatie 6, fluorescentie microscopie lokalisatie 7, evenals individuele molecuul Förster Resonance Energy Transfer (smFRET) 8, 9. Aangezien al vroeg FRET is genoemd moleculair liniaal, door de afstand afhankelijkheid van de lengteschaal van biomacromoleculen 10.
Een bijzonder interessante toepassing van smFRET is de afstand informatie van smFRET metingen gebruiken om afleiden structurele informatie 11, 12, 13, 14, 15 </sup>, 16, 17, 18, 19, 20, 21, 22, 23. Vanwege de hoge tijdsresolutie van smFRET kan de positie van de beweegbare delen van een eiwitstructuur worden gelokaliseerd. Echter, om kwantitatieve informatie smFRET data belangrijk correctieparameters de kleurstofmoleculen extraheren moet tijdens de meting 24 te bepalen. Met deze correctiefactoren, kan de FRET efficiëntie E FRET worden berekend met de formule
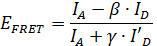 ,
,
waar ik A pt I D </sub> de fluorescentie-intensiteit van de donor en de acceptor molecule zijn (zie figuur 2). De β-factor is goed voor cross-talk, het lekken van donor-emissie in de acceptor kanaal en wordt berekend door

waarin I 'en A Ik zou de fluorescentie-intensiteit van de donor en de acceptor molecule na bleken door het acceptormolecuul.
Het γ-factor corrigeert het verschil in de relatieve detectie-efficiëntie van de twee kanalen en de verschillen in de fluorescentie kwantumopbrengst van de donor en de acceptor kleurstof. Het wordt berekend op basis van elk individu tijd spoor door
<p class="jove_content" fo:keep-together.within-page="1" fo:text-align="center" style = "text-align: center;">
Merk op, dat deze beschrijving verwaarloost directe excitatie van de acceptor molecule, die soms belangrijk wordt en zou moeten worden gecorrigeerd ook. Voor het vaststellen van deze correctiefactoren is het nuttig om zowel de donor als de acceptor wekken in een afwisselend schema 25 om onderscheid te maken tussen foto-fysische veranderingen en structuurdynamica.
Om niet alleen kwantitatieve smFRET efficiëntie maar ook kwantitatieve structurele informatie te verkrijgen, de Nano-Positioning System (NPS) werd in 2008 26. De naam werd gekozen op basis van de overeenkomsten met de satelliet-gebaseerde Global Positioning System (GPS). De NPS is een hybride techniek combineert smFRET en röntgenkristallografie gegevens voor het lokaliseren van onbekende kleurstof posities in Biomacromoleculaire complexen. de crystal structuur fungeert als referentieframe en de smFRET resultaten worden gebruikt om informatie afstand tussen een onbekende fluorofoor positie (antenne) en staat bekend uit de kristalstructuur (satelliet) te verkrijgen. In opeenvolgende experimenten werd de afstand tussen de antenne en meerdere satellieten worden gemeten en de positie van de antenne wordt bepaald met behulp van een statistisch rigoureuze analyseschema Bayesian parameterschatting. Daardoor is niet alleen de meest waarschijnlijke positie van de antenne berekend, maar de volledige 3D onzekerheid distributie, de zogenaamde posterior, gevisualiseerd door geloofwaardige volumes. Bovendien NPS uitgebreid, teneinde de analyse van complete netwerken smFRET 27.
De NPS is gebruikt om een aantal belangrijke vragen in eukaryote transcriptie, namelijk tijdens het stroomopwaartse DNA, de niet-matrijs-DNA en de ontluikende mRNA in het RNA polymerase II elongatie co lossenmplex 12, 28, blijkt ook het effect van transcriptie initiatie factoren 26 en de dynamische architectuur van een open-promotor complex 29. Bovendien, de NPS werd gebruikt om de structuur van de Archaea RNA Polymerase geopend complex 30 met name de positie van transcriptie initiatie factor TFE, die competitief aan dezelfde plaats als transcriptiefactor elongatie factor Spt4 / 5 31 helderen.
Sindsdien zijn er een aantal smFRET gebaseerd structurele aanpak is gepubliceerd op 15, 18, 21, 23. Bij het vergelijken van verschillende smFRET gebaseerd structurele methoden, wordt duidelijk dat de schijnbare nauwkeurigheid van de werkwijze sterk afhankelijk van de bepaalde keuze van kleurstof modellen. Men moet er rekening mee datkleurstofmoleculen kunnen verschillende ruimtelijke en oriënterende gedrag vertonen afhankelijk van de lokale omgeving.
Hiertoe werd Fast-NPS 32 geïntroduceerd. Fast-NPS maakt gebruik van een geavanceerde sampling algoritme het verminderen van de rekentijden drastisch. Bovendien, Fast-NPS maakt het mogelijk om een structurele analyse uit te voeren en voor elke kleurstof molecule kan de gebruiker kiezen uit een reeks van vijf verschillende kleurstof modellen die volgende zal worden beschreven. De meest conservatieve model, de zogenaamde klassieke, gaat ervan uit dat de kleurstof neemt slechts één, maar onbekende, positie. Bij deze stand kan de fluorofoor vrij draaien binnen een kegel, waarvan de grootte wordt bepaald door zijn respectieve (tijdsafhankelijke) fluorescentie anisotropie. De oriëntatie van de conus is onbekend, wat leidt tot grote onzekerheden bij het converteren van gemeten smFRET efficiëntie in afstanden. In dit opzicht is het model conservatief, omdat het leidt tot de kleinste precisie vergeleken met andere kleurstoffen modusls. Alleen voor zeer korte afstanden moet de aannames van het klassieke model leiden tot een merkbaar onjuiste positiebepaling. Voor typische smFRET waarden wordt de juiste positie steeds ingesloten in het relatief grote volume geloofwaardig.
Aangezien een hogere nauwkeurigheid gewenst is, is het belangrijk om te ontwikkelen en te testen alternatieve modellen kleurstof, die kan bijdragen tot de nauwkeurigheid te verbeteren. Als de kleurstof veel sneller dan zijn inherente fluorescentie levensduur roteert, kan de zogenaamde iso model worden toegepast. Hier is de oriëntatie factor K2 (nodig voor het berekenen van de karakteristieke isotrope Förster radius  ) Is ingesteld op 2/3. Dientengevolge, de berekende geloofwaardige volumes bijna twee orden van grootte kleiner in vergelijking met die in de klassieke model 32. In het geval dat de fluorofoor in een omgeving die niet alleen snel maakt reorientatie, maar daarnaast snelle beweging over haar toegankelijke volume moet de meanpos-iso-model worden gebruikt. In dit model, de kleurstof effectief neemt slechts een gemiddelde positie, waarbij de ruimtelijke middeling wordt verklaard door een polynoom afstand conversie 15. Dit model is van toepassing als bijvoorbeeld de (algemeen hydrofobe) kleurstof wordt een hydrofiel gebied, bijvoorbeeld het DNA bevestigd. Toepassing van de meanpos iso-model leidt tot een verdere verkleining van de geloofwaardige volumes met een factor van ongeveer twee. Echter, kan een kleurstof gebonden aan een eiwit reversibel binden aan verscheidene hydrofobe vlekken in het sterisch toegankelijke volume (AV). Een fluorofoor dat onmiddellijk schakelt tussen deze regio's, maar binnen één regio ondergaat vrije rotatie en snelle gelokaliseerde beweging wordt het best beschreven door de var-meanpos-iso-model. Voor een vergelijkbare situatie waarbij de kleurstof niet vrij de var-meanpos model geldt roteren. meer d IJZE over deze modellen kunt u vinden in onze recente publicatie 32.
) Is ingesteld op 2/3. Dientengevolge, de berekende geloofwaardige volumes bijna twee orden van grootte kleiner in vergelijking met die in de klassieke model 32. In het geval dat de fluorofoor in een omgeving die niet alleen snel maakt reorientatie, maar daarnaast snelle beweging over haar toegankelijke volume moet de meanpos-iso-model worden gebruikt. In dit model, de kleurstof effectief neemt slechts een gemiddelde positie, waarbij de ruimtelijke middeling wordt verklaard door een polynoom afstand conversie 15. Dit model is van toepassing als bijvoorbeeld de (algemeen hydrofobe) kleurstof wordt een hydrofiel gebied, bijvoorbeeld het DNA bevestigd. Toepassing van de meanpos iso-model leidt tot een verdere verkleining van de geloofwaardige volumes met een factor van ongeveer twee. Echter, kan een kleurstof gebonden aan een eiwit reversibel binden aan verscheidene hydrofobe vlekken in het sterisch toegankelijke volume (AV). Een fluorofoor dat onmiddellijk schakelt tussen deze regio's, maar binnen één regio ondergaat vrije rotatie en snelle gelokaliseerde beweging wordt het best beschreven door de var-meanpos-iso-model. Voor een vergelijkbare situatie waarbij de kleurstof niet vrij de var-meanpos model geldt roteren. meer d IJZE over deze modellen kunt u vinden in onze recente publicatie 32.
Deze modellen bieden een uitgebreid repertoire om specifiek rekening te houden met de verschillende omgevingen een kleurstof tegenkomen en de toepassing ervan optimaliseert wijselijk de lokalisatie precisie. Fast-NPS elke kleurstof molecuul gekoppeld aan een bepaalde positie kan worden toegewezen aan een specifiek voertuig, zodat FRET-partners mogen verschillende modellen. Dit maakt het mogelijk onbeperkt en close-to-natuur modellering. Het is echter belangrijk dat men voert strikt statistische proeven uit teneinde de door de uiteindelijke modellencombinatie resultaat nog steeds in overeenstemming met de experimentele gegevens. Deze tests zijn in de Fast-NPS software.
Om Fast-NPS toepassing op experimentele gegevens van de meting (alleen) drie invoerparameters vereist. Ten eerste, de kleurstof-pair specifieke isotrope Förster radii (/54782/54782eq5.jpg "/>) Moeten worden bepaald. Daarom is de kwantumopbrengst (QY) van de donorkleurstof, de donor fluorescentie-emissiespectra en de acceptor absorptie spectra te meten nodig. Deze metingen kunnen worden uitgevoerd in bulk, met een standaard spectrometer en een standaard fluorescentie spectrometer. Voor elk paar, het R 0 wordt vervolgens berekend met de freeware PhotochemCAD en kan worden gebruikt in de NPS analyse. Ook de (Tijdsgeresolveerde) fluorescentie anisotropie van de kleurstofmoleculen moeten worden verkregen met een polarisatie (en tijd) gevoelige fluorescentie spectrometer. de belangrijkste invoerparameters voor Fast-NPS zijn smFRET rendementen gemeten op een enkel-molecuul fluorescentie microscopie setup, zoals een totale interne reflectie fluorescentie microscoop (TIRFM) .
Hier presenteren we een stap-voor-stap protocol voor het verkrijgen smFRET databank toepassen Fast-NPS (figuur 1).
