The present protocol describes the sequential steps for establishing a murine model of ocular surface inflammation. The protocols aim to show how to apply therapeutics locally, obtain ocular exudates, and excise associated accessory organs such as healthy and inflamed eyelids (Figure 2), the cornea, and the conjunctiva. Attention must be paid when the upper eyelids are dissected for the isolation of the conjunctiva, and it must be stored in 1x PBS during the dissection of the cornea. This will prevent the drying of the conjunctiva, which can be used for histological, pharmacokinetic, and gene expression studies.
OVA and saline were applied topically for 7 consecutive days following the abovementioned protocol. Mice challenged topically with saline showed a healthy ocular surface with wide-open eyes and a regular blinking pattern. However, ocular inflammation was instigated in immunized C57BL/6J mice challenged with OVA. The instillation of OVA solution on the ocular surface caused itching, and not pain, for the first 2 hours after instillation. Exudate and eyelid eczema was observed only during the last 3 days of the challenge phase. No pain was evident, judging the behavior of the animals. Therefore, a moderate level of stress for the mice over a short period of time was assumed. The daily OVA challenge triggered clinical manifestations like abundant ocular discharge, chemosis, and narrow opening of the eyes. In addition, the upper and lower eyelids frequently adhered to each other, impairing the essential function of blinking (Figure 2). Strict interruption criteria (Supplementary File) were followed for this model and were approved by the local ethical board for animal experimentation. The immediate interruption was carried out if one mouse reached 15 points at any given time.
Supplementary File. Please click here to download this File.
After euthanasia, the ocular organs were excised and were observed at higher magnification. The excised eyelids under a microscope showed large occlusions plugging the orifices of the MG and edema, in stark contrast to the healthy eyelids showing small plugs of the gland lining the eyelid (Figure 3).
Further examination of the gland's infrared transillumination reveals the racemic appearance of the acini of the MG. This allows for quantifying the visible Meibomian glands (indicated in red). The eyelids from saline-challenged mice showed the round acini forming the MG. In comparison, the application of OVA induced the destruction and loss of some MG in mice with allergic eye disease (AED, Figure 4). The histological analysis of the eyelids displayed dilated MG compared to naïve mice administered with only saline for 7 days (Figure 5), allowing the accumulation of neutrophils producing aggNETs that finally obstruct the orifices of the gland.
Separating the cornea from the eye's sclera can be cumbersome due to the slippery mucous surface of the eye globe. Incubating the eyeball on dry ice in a Petri dish for 3 min allows for fixing the eye, making a small incision at the limbus, and dissecting the cornea (Figure 6).
The steps enlisted in the protocol section facilitated the collection of the cornea and conjunctiva. In addition, the local administration of OVA inflicted severe inflammation to the conjunctiva, with a hyperaemic appearance (Figure 7).
The analysis of ocular exudates reveals molecular mechanisms in the development of MGD. Cytokine and chemokine quantification as described showed elevated levels of the major chemokine facilitating neutrophil extravasation, phagocytosis, and degranulation (CXCL-1) in the supernatants from mice with AED8. In addition, the prime mediator of the acute phase response and neutrophil production, IL-6, was also significantly elevated in mice with AED. On the other hand, the concentration of IL-10, an anti-inflammatory cytokine, showed no significant changes in both naïve and AED mice (Figure 8).

Figure 1: Excision of the facial tissue surrounding the orbital region. The incision trail is indicated in red. This allows for excising the ocular organs and investigating the influence and effects of various topically administered therapeutics on accessory ocular organs such as the conjunctiva and cornea. Please click here to view a larger version of this figure.
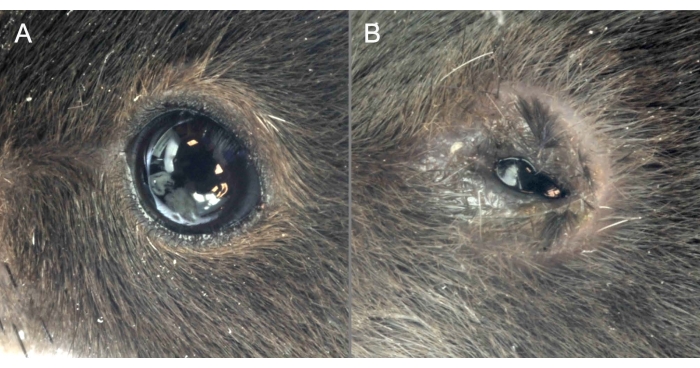
Figure 2: OVA administration causes clinical manifestations of MGD. (A) Mice administered saline show a healthy ocular surface with eyes wide open. (B) OVA application induces severe ocular surface inflammation, the narrow opening of the eyes, and signs of chemosis. Please click here to view a larger version of this figure.
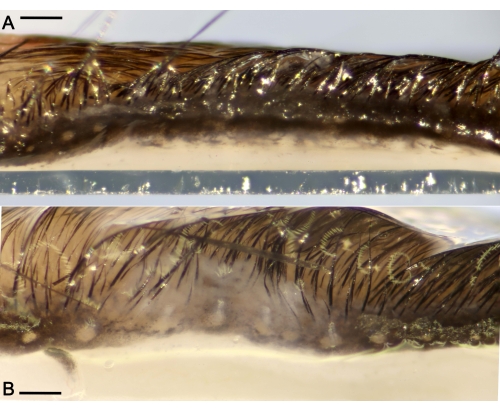
Figure 3: Ductal plugs obstructing the Meibomian glands (MG) located along the eyelids. (A) Representative macro photograph of a healthy eyelid without excessive ocular discharge from a mouse administered saline for 7 days only. (B) Eyelid showing large ductal occlusions of the MG and edema after the challenge period. Scale bar = 300 µm. Please click here to view a larger version of this figure.
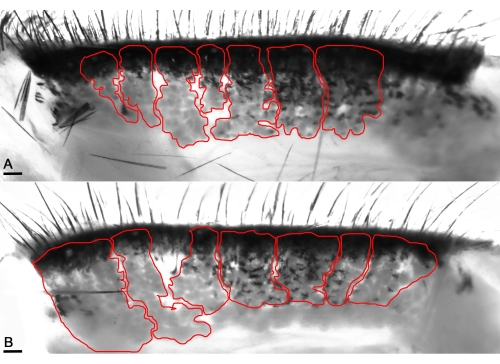
Figure 4: Transilluminating macrophotography enables the visualization of the MG's acini (red lines). (A) Enhancing infrared levels visualizes the healthy acini of naïve mice and unveils the distinct pocket-shaped acini in the healthy eyelids. (B) The acini appear thick in the mice's affected eyelids with OVA application. Scale bar = 100 µm. Please click here to view a larger version of this figure.
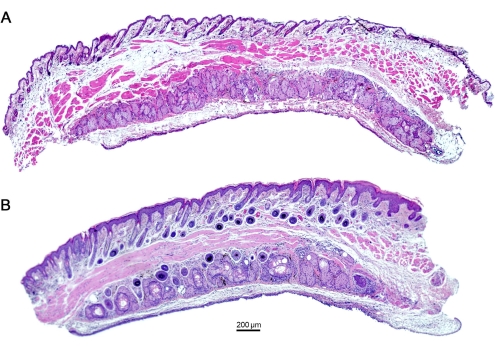
Figure 5: Histological analysis of the eyelids shows dilated ducts of MG in mice with repeated OVA insult for 7 days. (A) Naïve mouse eyelids show the absence of any dilated ducts, depicting a healthy functioning ocular organ (B) in contrast to mice with OVA challenge. Scale bar = 200 µm. Please click here to view a larger version of this figure.
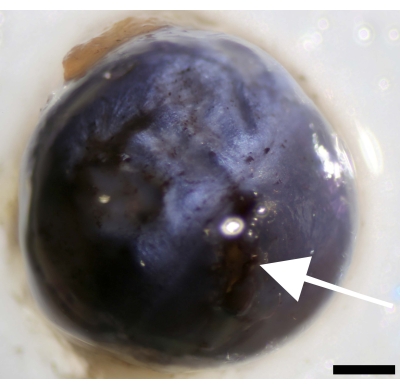
Figure 6: Incision at the cornea next to the limbus and separation of the cornea from the sclera. Image of the eyeball showing the point of incision (indicated by white arrow). Scale bar = 500 µm. Please click here to view a larger version of this figure.
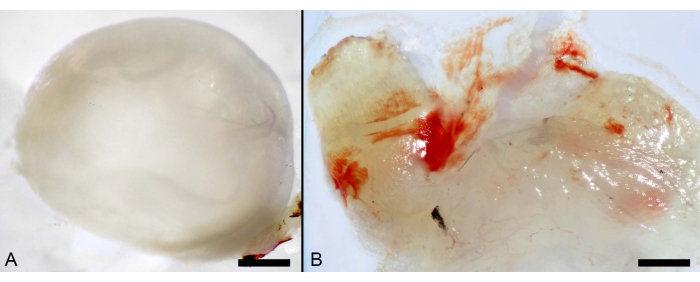
Figure 7: Local administration of OVA. (A) Macro photographs of murine ocular organs showing the cornea. Scale bar: 600 µm. (B) Inflamed conjunctiva from mice challenged with OVA. Scale bar: 100 µm. Please click here to view a larger version of this figure.
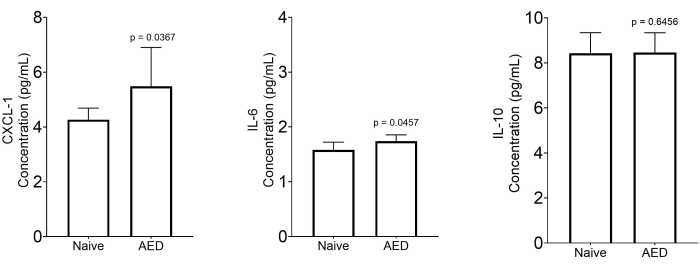
Figure 8: Measurement of inflammatory markers in the collected ocular exudates. Quantitative analysis of cytokines and chemokines in the supernatant of centrifuged ocular exudate of naïve mice (n = 7) and mice with MGD (n = 8). Data are expressed as median with a 5%-95% range. Statistical significance was calculated by using the two-tailed Mann-Whitney U-test. Please click here to view a larger version of this figure.
