Using this technique, the total cell count of the naïve dissociated lungs (only the left lobes were used for the representative data) was between 27.3 x 106 to 71.1 x 106 cells/mL. After gating on size and gating out doublets and dead cells (gating scheme in Figure 2), the leukocyte counts ranged from 6.9 x 106 to 13.5 x 106 cells/mL. Circulating leukocytes that remain trapped even after perfusion to clear the lungs constituted approximately 4% to 13% of the live cells in the experiment. While all leukocytes in the dissociated lung stain for CD45 on the PE-Cy7, only the circulating leukocytes are double stained with the CD45 antibody on both fluorophores, PE-Cy7 and Pac blue. Despite standardizing the process of perfusion, we found that about 24% to 70% of the total leukocytes in the dissociated lung belonged to the circulating leukocyte pool. The mean leukocyte counts without accounting for the intravascular labeling was 10.2 x 106 cells/mL, while the corrected value was 4.93 x 106 cells/mL (Figure 3A). Therefore, failing to differentiate residual intravascular leukocytes that persist after perfusion results in a significant overestimate the number of pulmonary leukocytes. For this experiment, animals were 8-12 weeks of age and weighed 25-30 g at the time of dissection. Dose adjustments may need to be made for extremes of ages and weight. Other useful controls include using an intravascular sample to demonstrate good labeling of intravascular leukocytes and a sample from an unlabeled tissue (such as a lymph node) to exclude the possibility to diffuse labeling of all tissues by transmigration of labeled leukocytes from the blood.
Since research questions often involve circadian sampling, we further broke down the data by time of day (for the animal) at which the lung harvest was performed. By circadian convention ZT0 refers to the time when lights turn on. We used dawn (ZT23: onset of rest phase) and dusk (ZT11; onset of active phase) as the two time points. We note that the proportion of the intraparenchymal and the residual intravascular leukocytes varied by the time of day at dissection (Figure 3B). In this experiment, the mice were maintained in reverse light-dark (LD) cycles using light-controlled circadian boxes, so that ZT11 and ZT23 mice were in reverse phases at the same conventional time and could be harvested simultaneously. Thus, lungs from both groups were labeled, harvested and digested at the same time of the day and therefore these differences in Figure 3 cannot be attributed to differences in processing.
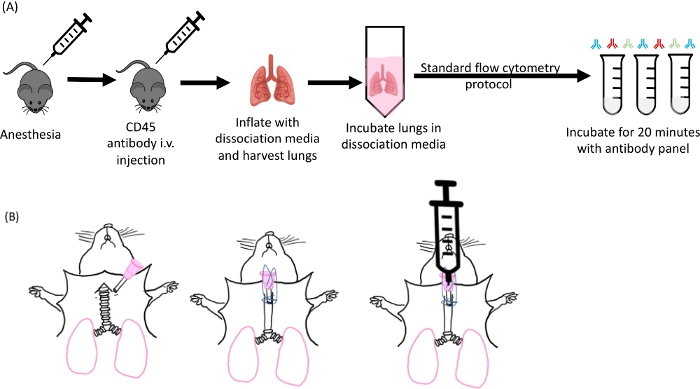
Figure 1: Schematic of lung inflation and digestion. (A) Overall experimental design starting from anesthetizing of the mice to the antibody staining step. (B) Image of the cannula insertion step of the lung harvest. Please click here to view a larger version of this figure.
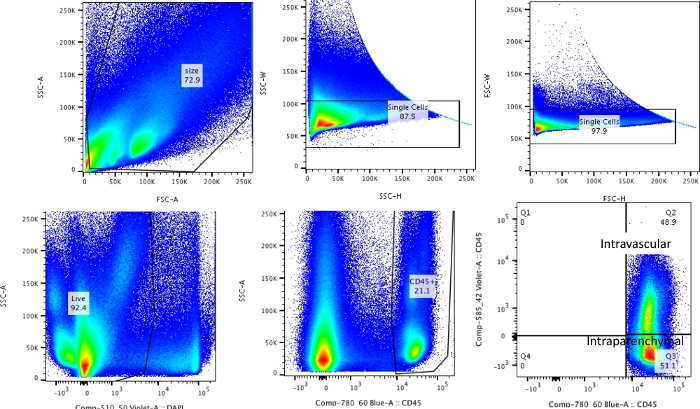
Figure 2: Gating strategy. Gating strategy used for differentiating intravascular leukocytes from leukocytes that are truly resident in the lungs. Please click here to view a larger version of this figure.
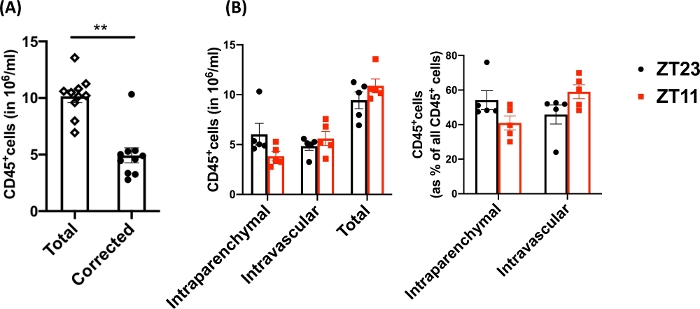
Figure 3: Comparison of intraparenchymal and intravascular leukocytes. (A) Leukocyte counts (in 106 cell/mL) using traditional digestion and labeling compared with the counts corrected for residual intravascular leukocytes that were not cleared out of the lungs. P<0.0001 by Student’s t-test. Data represented as mean±SEM. (B) Leukocyte numbers and percentage from lungs harvested at dawn (ZT23) or dusk (ZT11). (right panel) 2-way ANOVA, p >0.05 for both time of harvest and compartment labeled, but p<0.05 for interaction. Data represented as mean±SEM. Please click here to view a larger version of this figure.
