The temperature-controlled arena (Figure 1A) consists of three copper tiles whose temperature can be individually controlled through a programmable circuit. Each copper tile possesses a temperature sensor that gives feedback to the programmable circuit. The circuit activates a power supply to increase the temperature of each tile. Passive thermoelectric elements act as constant heating elements to maintain the desired temperature, while a heat sink cooled by a fan provides constant cooling. The magnitude of temperature change determines the speed of the process in a non-linear manner. An increase of 2 °C requires only 0.1 s, and an increase of 18 °C requires 4 s. A screen connected to the programmable circuit (Figure 1C) informs the user of the temperature measured by the temperature sensors in each of the tiles. The copper tiles are surrounded by an aluminum ring constantly heated to 50 °C (Figure 1B and 1C) by semiconductors around the periphery. This ring forms the edges of the Fly Arena (Figure 1C), the area in which flies are to be placed. The Fly Arena is covered by a siliconized glass cover (Figure 1A and 1C), which provides a 3 mm high space which ensures that flies can walk but not fly. Next to the Fly Arena are two red LEDs (Figure 1C) that can be programmed to mark different experimental phases. For example, for the results shown in Figure 2A, each LED is associated with a different temperature, while in Figure 2B, each LED indicates 60 s. The FlySteps software can register when each of the indicative LEDs is on, and the researcher can then use this information to automatically determine the experimental phases based on temperature or time.
The temperature-controlled arena can be used to compare the behavioral response of flies from different genetic backgrounds to dynamic temperature changes. For example, flies from different species can be exposed to gradually increasing temperatures (Figure 3) to compare differences in thermal performance. The speed of all species increases as temperature increases until reaching a point of maximum performance, after which it decayed and perished. However, each species has a particular response curve with specific maximum response speeds and thermal tolerances. Previous reports have shown that Drosophila from different species differ among developmental timing, longevity, fecundity, body dimensions, sexual communication, and temperature tolerance3,6,7,8,22. Thus, our description of species-specific locomotion in a temperature gradient adds to this body of work.
The temperature-controlled arena can also be used to explore the response to conditioning experiments based on temperature. The simplest form of this approach is an operant conditioning paradigm in which flies are trained to prefer one side of the arena over the other, by warming up the side that will be avoided23,24,25. We exposed individual flies to 40 °C in the middle and one of the side tiles, while leaving the other side tile at a comfortable 22 °C (Figure 4). Wild-type flies quickly stopped moving along the arena and remained in the comfortable location. In contrast, the classic memory mutant Dunce kept exploring the arena and spent less time than controls in the comfortable location. The differences between performance of the wild-type flies and Dunce mutants became larger when all tiles were set to 22 °C and comparisons were made between the treatment groups. Dunce mutants also showed greater differences between training and test phases in comparison to the wild-type flies (Figure 4). These results suggest an effect of memory on remaining in the comfortable location.
Combinations of temperature and location are also useful to understand the function of different temperature receptors during dynamic temperature changes. We exposed individual D. melanogaster Gr28b.d and TrpA1GAL4 mutants to increasing temperatures (2 °C increase every 60 s) while providing a comfortable location at 22 °C (Figure 5). The comfortable location shifted from left to right, and vice versa, per iteration. Results show that the periphery temperature receptor Gr28b.d mutants behave as the control, as they spend more time in the comfortable location as temperature increases. However, brain temperature receptor TrpA1GAL4 mutants are not affected by increasing temperatures and do not change their locations in the arena. The increases and decrease in the curve of TrpA1GAL4 mutants show the effect in flies that were already sitting in the comfortable location before it became comfortable and remained there during that phase. The consistency of peaks and valleys of the curve of TrpA1GAL4 suggest that these flies remained still for most of the experiment; hence, they were constantly counted when their location was the one considered comfortable. This conclusion was confirmed by visual inspection of the recorded videos. These results support previous physiological reports suggesting that periphery perception of fast and large changes does not depend on Gr28b.d17 and that flies possess a main central mechanism to sense temperature based on TrpA114,21.
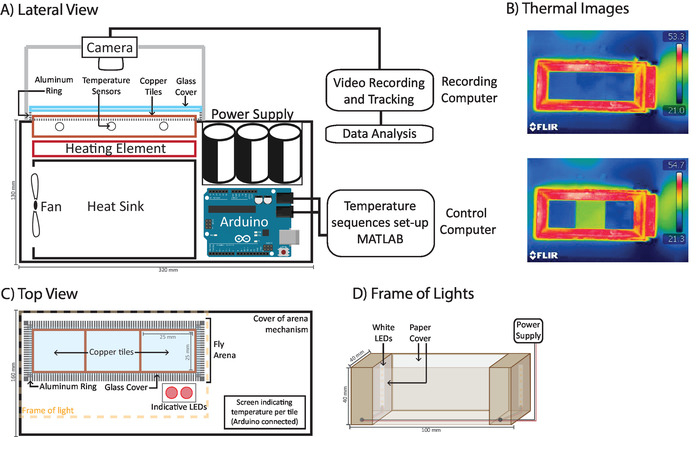
Figure 1: Diagram of temperature controlled-arena. (A) A lateral view of the temperature-controlled arena. A programmable circuit connects a power supply and temperature sensors to heating elements under copper tiles to control their temperature. Tiles are constantly cooled down through a heat sink connected to a fan. A heated aluminum ring over which a glass cover rests surrounds the tiles. (B) Thermal imaging showing the tiles set at 24 °C (top) and side tiles at 24 °C with a middle tile at 30 °C (bottom). (C) A top view of the arena. A camera records the copper tiles, aluminum ring, and red LEDs, then automatically determines experimental phases. A screen in the corner of the box, not recorded by the camera, displays the current tile temperature. (D) Ring of light: two warm white LED strips inside a wooden box covered in white paper ensure constant and symmetric illumination of the whole arena. Please click here to view a larger version of this figure.
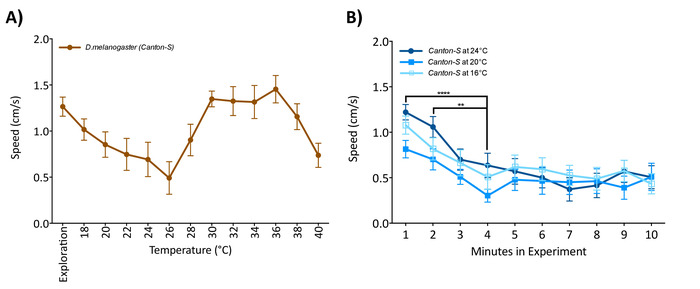
Figure 2: Flies must acclimate to the arena before starting the temperature protocol. (A) Single male flies were introduced to the arena and allowed to explore at a constant 16 °C for 1 min, after which the temperature started increasing. (B) Single flies exposed to 16 °C, 20 °C, or 24 °C (no group differences; two-way ANOVA F (2,570) = 4.156, p = 0.162) have a higher locomotion at the beginning of the experiment than after 5 min (two-way RM ANOVA F (9,570) = 7.803, p < 0.0001). Data are mean and standard error of the mean (± SEM) of 20 virgin female flies 5 to 7 days old tested over multiple days. Asterisk indicates significant difference among groups (****p < 0.0001; Tukey's multiple comparison test, p = 0.05). Please click here to view a larger version of this figure.
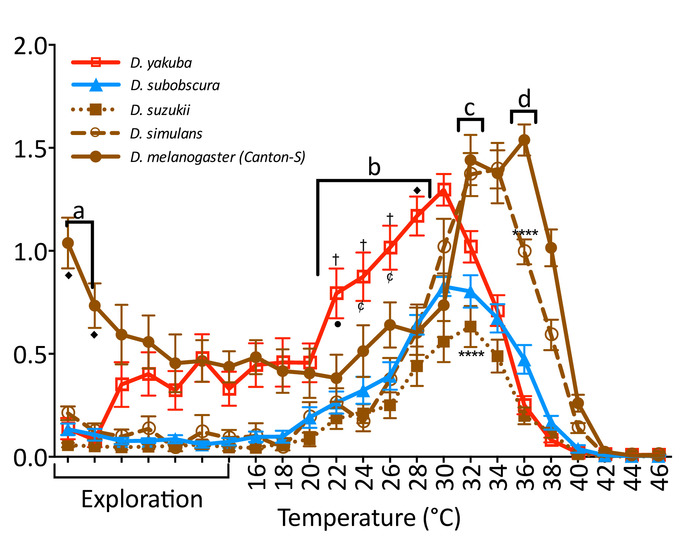
Figure 3: Locomotion of 5 Drosophila species exposed to gradually increasing temperatures. Individual male flies from temperate (blue), tropical (red), and cosmopolitan (brown) Drosophila species were exposed to an increasing temperature gradient (2 °C every 60 s) between 16 and 46 °C. The first 7 min were constantly at 22 °C to allow flies to explore the arena. Species were significantly different (two-way RM ANOVA F(4,70) = 28.46, p < 0.001). (a) D. melanogaster (brown; filled circles) was faster when introduced to the arena. (b) D. yakuba (red; empty squares) was faster as temperature increased. (c) D. suzukii (brown; filled square) was slower than the other cosmopolitan flies at its maximum performance point. (d) D. simulans (brown; empty circles) was in decay at the maximum point of D. melanogaster. Each point represents the mean (± SEM) of 15 male flies 5 to 7 days old tested over several days. Significance indicated by symbols (♦ = difference from all, p < 0.0001; †= difference from all except D. melanogaster, p < 0.0001; • = difference from D. melanogaster, p < 0.01; ¢ = difference from D. melanogaster, p < 0.001; **** = difference between named groups, p < 0.0001; Tukey's multiple comparison test, p = 0.05). Please click here to view a larger version of this figure.
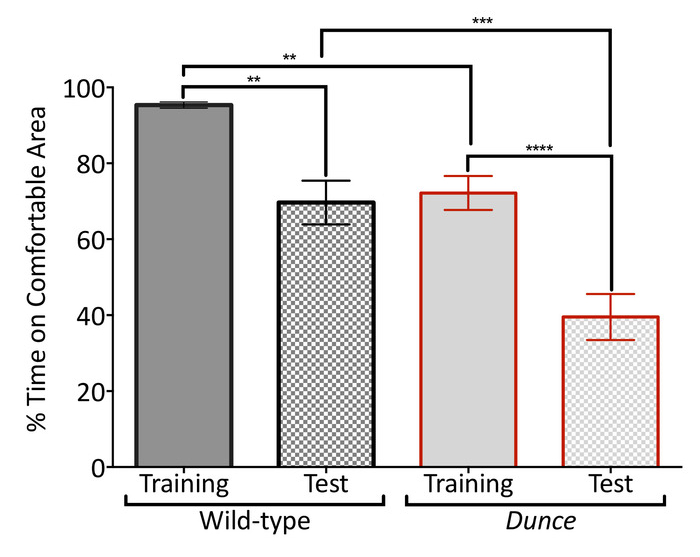
Figure 4: The temperature-controlled arena can be used for operant conditioning. D. melanogaster Canton-S strain (wild-type; black border) and dnc1 (Dunce; red border) mutants were trained to prefer a lateral tile at 22 °C after warming the middle and opposite lateral tiles to 40 °C for 4 min (training, no pattern). Memory of the heated areas is then tested by setting all tiles to 22 °C (test; grid pattern). Flies were conditioned to prefer tiles on the left in half of the experiments, then tiles on the right in the other half. The percentage of total time inside the tile at 22 °C during training and testing was measured to compare performances. Groups were significantly different (one-way ANOVA F(3,76) = 23.23, p < 0.0001), with Dunce performing worse than wild-type overall. Data are mean (± SEM) of 20 virgin female flies 5 to 7 days old tested over several days. Asterisks indicate significance difference among groups (****p > 0.0001; ***p > 0.001; **p > 0.01; Tukey's multiple comparison test, p = 0.05) Please click here to view a larger version of this figure.
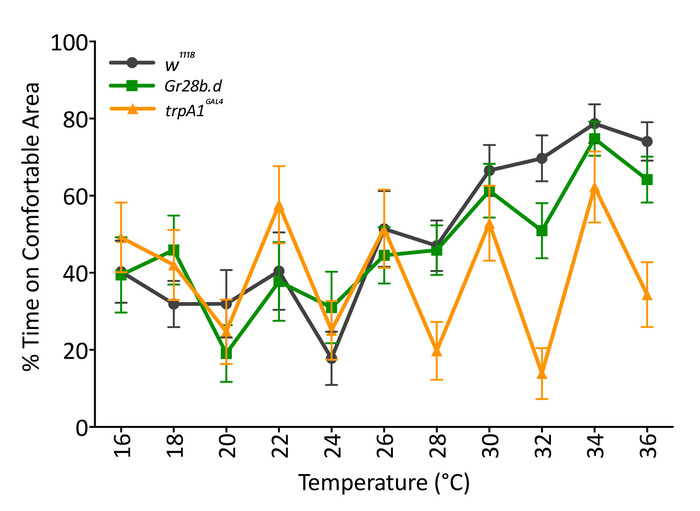
Figure 5: Response of temperature mutants to increasing temperature when a comfortable location is provided. Temperature mutants Gr28b.d (green; squares) respond as controls (w1118, black; circles) by increasing the percentage of time in the comfortable area as temperature increases (two-way RM ANOVA F (1,38) = 0.5107, p = 0.479). TrpA1GAL4 mutants (yellow; triangles) are different from controls (w1118, black), as they do not increase the time in the comfortable area as temperature increases (two-way RM ANOVA F (1,38) = 1.670, p = 0.019). Data are mean (± SEM) of 20 male flies 5 to 7 days old tested over several days. TrpA1GAL4 is significantly different from Gr28b.d and the control (p < 0.05; Tukey's multiple comparison test, p = 0.05). Please click here to view a larger version of this figure.
