1. Animal culture
- Maintain Clytia polyp colonies on microscope slides and medusae in artificial sea water (ASW) at 18 °C in a zebrafish system, with 2 L zebrafish tanks for polyp colonies and custom-made 5 L pseudo-kreisel tanks for medusae (Supplemental Figure 1)51. ASW consists of 4% Instant Ocean in deionized (DI) H2O.
- Feed the animals daily with 2-3-day-old artemia as described51.
NOTE: Wound healing imaging is easier if the animals have not been recently fed, as there is less debris released from the gut into the field of view. - Collect baby medusae from the established polyp colonies as needed by placing colonies in a 2 L beaker filled with 1 L of ASW overnight. Use 2-3-week-old female medusae for all wound healing experiments. Propagation of Clytia has been described in detail elsewhere51.
2. Wounding
- Creating microwounds within and between cells (20-500 µm2)
- Create a modified transfer pipette by cutting the tip with scissors to make a larger opening (0.5-0.7 cm diameter).
NOTE: The opening in the pipette should be wide enough to avoid any damage to the animal. - Using the modified transfer pipette, place the medusa on a depression slide with the medusa exumbrella facing up, with just enough ASW to cover the animal.
- Place a coverslip over the animal and image immediately (see below for description of imaging). The coverslip compresses the mesoglea, and the rebound of the compressed tissue creates a force that pushes the cells slightly apart52. This immediately appears as gaps between each cell and damage within some cells (Figure 1B,B', Figure 2, and Figure 3A-C).
- Create a modified transfer pipette by cutting the tip with scissors to make a larger opening (0.5-0.7 cm diameter).
- Creating small epithelial wounds (0.02-0.125 mm2)
- Using a modified transfer pipette (as above), place the medusa on a depression slide with the medusa exumbrella facing up.
- Using a 200 µL pipette tip, gently scratch the surface of the medusa. Gentle scratching can also create rips in the basement membrane, which are readily apparent22. Cover the animal with a coverslip for imaging. Alternatively, placement of the coverslip is sometimes sufficient to create small epithelial wounds even without scratching (Figure 1C,C', Figure 2, and Figure 3A-C).
NOTE: Do not press down when scratching the surface of the medusa, as this damages the ECM and creates an irregular surface — epithelial cells migrating on an irregular surface are more difficult to keep in focus.
- Creating large epithelial wounds (0.5-0.9 mm2)
- Make a microinjection needle using a micropipette puller and glass capillary tube (step 5.2). Place the empty microinjection needle into a microinjector holder affixed to a micromanipulator. Cut the tip of the needle so that the opening is approximately 20-40 µm.
NOTE: Cut needles for large epithelial wounds can be stored and reused to increase consistency between experiments. - Set the hold pressure on the microinjector to zero, and set the eject pressure to approximately 20 PSI. Set the microinjector to deliver a 2 s pulse of air.
NOTE: The eject pressure may need to be adjusted based on the diameter of the needle opening (i.e., smaller tips will use higher pressure, while larger tips will use lower pressure). - Place the medusa with the exumbrella facing up on a depression slide on the stage of a dissecting scope, with just enough ASW to cover the animal. Using the micromanipulator, adjust the microinjection needle tip so that it is just above the water. To do this, carefully dip the tip into water (water may enter the pipette tip), then retract it so that it is close to the medusa's epithelial surface.
NOTE: The tip should be positioned over one quadrant of the medusa. The medusa's radial canals divide the medusa bell into four distinct quadrants. Targeting a quadrant will result in cleaner imaging, as the gonads and the radial canals are excluded from the wound area. - Pulse air by pressing start on the injector. Repeat the pulse in the same spot two to four times, depending on the width of the tip. Larger tips require fewer pulses.
NOTE: An indent in the water/medusa caused by the pulse of air should be visible. - Cover the wounded animal with a coverslip for imaging large wounds (Figure 1D,D').
- Follow the steps below (section 3) for imaging epithelial wound healing.
- Make a microinjection needle using a micropipette puller and glass capillary tube (step 5.2). Place the empty microinjection needle into a microinjector holder affixed to a micromanipulator. Cut the tip of the needle so that the opening is approximately 20-40 µm.
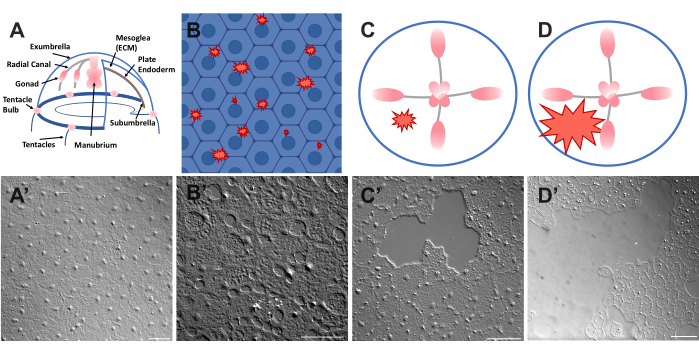
Figure 1: Intact and wounded exumbrella epithelial layer in Clytia medusa. (A) Cartoon graphic of the Clytia medusa body. (A') Intact medusa exumbrella epithelium viewed from above. (B) Cartoon of single-cell microwounds (red jagged shapes) with epithelial cells in blue. (B') Single cell microwounds. (C) Cartoon of a small epithelial wound (red jagged shape). (C') Small epithelial wound. (D) Cartoon of a large epithelial wound (red jagged shape). (D') Large epithelial wound. Images were all obtained using DIC microscopy. Scale bars in (A'–C'): 50 µm. Scale bar in (D'): 100 µm. Please click here to view a larger version of this figure.
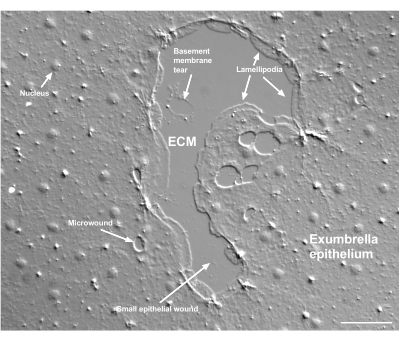
Figure 2: Multiple size wounds and a damaged basement membrane. A typical small exumbrella epithelial wound is shown, with labels indicating lamellipodia that form from marginal cells. In addition, microwounds within and between epithelial cells are seen. Note the small basement membrane tear in the upper portion of the wound. Movie 4 shows healing of this wound. Scale bar: 50 µm. Please click here to view a larger version of this figure.
3. Imaging epithelial wound healing
- Make sure that the microscope has been aligned for Köhler illumination53 and that it has been correctly set up for differential interfering contrast (DIC) microscopy54. Epithelial cells are almost invisible with standard optics (Figure 3D,E).
- Adjust the focus to the exumbrella. Although this is a thin layer, hexagonal cells should be clear.
NOTE: The exumbrella and subumbrella are separated by a thick mesoglea that is supported by vertical fibers. The subumbrellar cells are in the same focal plane as the radial canals. If initially focused on the subumbrellar layer, then adjust the focus slowly through the mesoglea and vertical fibers until finding the exumbrella. - Manually identify a wound to image. For large wounds, use a 10x objective. For smaller wounds and single-cell wounds, use a 20x objective.
- Start a program that collects images as a movie in real time or that collects a series of images at regular intervals. Monitor the progress to make sure that the wound area does not drift out of the field of view and that the cells of interest remain in focus.
- Single-cell wounds close within a minute; therefore, image their closure with a movie.
- To capture details of cell dynamics for small wounds, collect images approximately every 10 s. Closure of small wounds takes 20-50 min depending on size.
- Do not image the unsealed slides for more than 45 min, as evaporation of water from the slide over time leads to animal death and rupture of the cells.
- For longer observation, seal around the coverslip with petroleum jelly to reduce evaporation.
NOTE: Some medusa may pulse on the slide, which interferes with imaging. In this case, mounting animals in a 1:10 dilution of 1% Ethyl 3-aminobenzoate methanesulfonate (Tricaine), adjusted to pH 7.5, in ASW serves as an effective anesthetic and has no apparent effect on healing in a 1 h time frame. However, the animals will die if left for several hours in Tricaine.

Figure 3: Creating a small wound in the exumbrellar epithelium. (A) Gentle scratching of the exumbrella with a 200 µL pipette tip to create a small epithelial wound. (B) Placing the coverslip is sometimes sufficient to create small epithelial wounds. (C) Medusa mounted on a depression slide. (D) Small epithelial wound image without DIC optics and (E) with DIC optics. Scale bars: 50 µm Please click here to view a larger version of this figure.
4. Analysis
- Preparing image files
NOTE: To process the image files, use FIJI/ImageJ with updated BioFormat plugins.- Set the scale to the correct pixel per micron ratio before registering the image stack; Analyze > Set Scale. This is necessary for extracting actual size measurements in downstream analyses.
- Often, the animal drifts slightly on the microscope slide; therefore, to eliminate drift in movies, register the images using the FIJI plugin linear stack alignment with SIFT. Plugins > Registration > Linear Stack Alignment with SIFT.
- Save the registered stack as a .avi file. File > Save As > AVI… In the pop-up, set the frame rate (animated figures herein are set to 10 fps) and click OK. Use this output to perform wound healing analysis.
- Analysis of wound area
- Using the lasso tool in FIJI/ImageJ, outline the wound by tracing the cell edges. Measure the wound area that was just outlined with Command+M or CTRL+M.
- Repeat wound area measurement every 10 frames. The measurements from FIJI/ImageJ can then be plotted using Prism 9 (Figure 4).
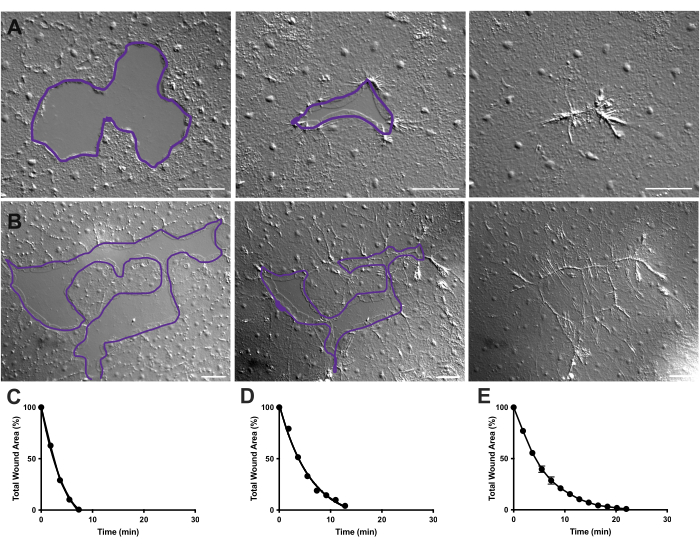
Figure 4: Analysis of wound area in small epithelial wounds. (A) Example of a small epithelial wound healing over 10 min. (B) Example of a different epithelial wound healing over 21 min. The purple outlines in A,B are comparable to the measurements of wound areas using the lasso tool in FIJI/ImageJ. (C) Normalized reduction of the wound area over time in A. (D) Normalized reduction of the wound area over time in B. (E) Average reduction of the wound area over time for 14 small wounds. n = 14. Error bars centered around mean ± SEM. Scale bars: 50 µm Please click here to view a larger version of this figure.
5. Mesogleal injections
- Creating injection dish
- Prepare polydimethylsiloxane (PDMS) by combining a PDMS base and curing agent, in a ratio of 10 parts base to 1 part curing agent by weight. Stir vigorously to fully mix the base and the curing agent.
- To remove bubbles, put the mixture in a vacuum chamber for 15 min. Pour the mixture into a 60 mm Petri dish with microcentrifuge tube caps to hold the mold in place. Immediately place the mold on tube caps at a 45° slant and tape in place. The mold is three stacked, offset glass slides glued together to create ridges in the final injection dish.
- Place the entire dish, mold, and mixture into an oven at 60 °C for 2 h to cure the elastomer. Remove the mold for a completed injection dish.
- Micropipette pulling
- Using a microelectrode puller, design a pulling program. Use a one-step program with high velocity. The heat is approximately the glass RAMP test result55,56. Check resulting micropipettes for long consistent tapers.
NOTE: Use thin wall glass borosilicate capillaries with a 1.0 mm outer diameter, 0.75 mm inner diameter, and 10 cm length.
- Using a microelectrode puller, design a pulling program. Use a one-step program with high velocity. The heat is approximately the glass RAMP test result55,56. Check resulting micropipettes for long consistent tapers.
- Injection of dyes and drugs
- Make a microinjection needle (as above).
- Backfill the microinjection needle using a long pipette tip with an excess volume of dye or drug for injection into the medusa.
NOTE: For Clytia, dimethyl sulfoxide (DMSO) should be kept at a <1:100 dilution with ASW, as higher DMSO concentrations impede wound healing. If injecting a clear solution, Fast Green FCF solution (1:100 dilution of 0.1% Fast Green FCF in ASW) can be added to visualize the injected liquid. - Using a modified transfer pipette as above, place a medusa with the subumbrella facing up into a PDMS injection dish with just enough ASW to cover the animal (Figure 5C). Place the dish on the stage of a dissecting scope.
NOTE: Limiting excess ASW prevents the medusa from swimming in the dish and allows more successful injections. - Focus on the microinjection needle tip and advance it into the water near the medusa. With the micromanipulator, press the needle into the dish until it bends and breaks. This tip opening is approximately 10-20 µm.
NOTE: This needle can be used repeatedly for the same dye/drug injections that day. It is recommended to use a fresh tip each day and for separate dyes/drugs. - Using the micromanipulator, insert the tip of the needle through the subumbrella into the mesoglea without puncturing the exumbrella.
NOTE: A creasing/folding of the epithelium will be noticeable. Once the needle is inserted into the medusa, the creasing/folding ceases. - On the microinjector, set the hold pressure to zero and ejection pressure to ≤20 PSI. Inject into one or two quadrants, filling each with a spot of dye or drug roughly 1/4 of the area of that quadrant.
NOTE: Depending on the size of the medusa, larger or smaller volumes are appropriate in single injection spots. Overfilling the medusa results in extreme damage to the epithelium and even death of the animal. - Depending on what dye or drug is being injected, animals are placed into a beaker of fresh ASW to allow for dye or drug diffusion and incubation.
- For imaging, mount the medusa onto a depression slide using a modified transfer pipette, positioning the animal so the exumbrella is facing up (Figure 5). Animals can be wounded at this stage to test the effect of an injected reagent.
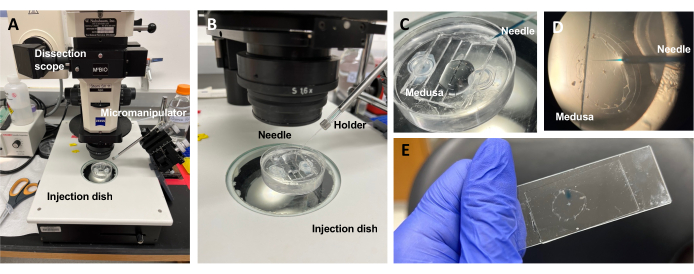
Figure 5: Injection setup for introducing dyes or drugs to the ECM. (A) Injection setup. (B) Close-up of the injection setup showing microinjection needle orientation (approximately 45° angle relative to the animal in the dish). (C) Close-up of the silicone injection dish with the medusa in a small amount of ASW for injection. (D) A microinjection needle loaded with Fast Green FCF entering the mesoglea of the medusa through the subumbrella. (E) Post-injection of Fast Green FCF in a mounted medusa. Please click here to view a larger version of this figure.
Following the protocols above, single-cell microwounds, small wounds, and large wounds were imaged. Registered stacks of image files were saved as .avi files.
In Movie 1, microwounds can be seen to close between and within cells (Figure 1 and Figure 2). Small lamellipodia are observed during closure, followed by contraction and healing. Debris is excluded and released into the water. Healing is completed in a minute or less.
In Movie 2 and 3, small wounds of different shapes heal through the formation of lamellipodia, extension of lamellipodial contacts, and spreading of cells at the wound margin, as previously described40 (Figure 1 and Figure 2). Cells in tiers behind the marginal cells do not participate in healing of wounds of this size nor is there collective cell migration. Rapid and progressive closure of epithelial gaps is followed by tissue contraction along the newly formed wound seam40. The normalized rate of healing of these two wounds, expressed as a percentage of the original area over time, is shown (Figure 4C,D). While there is some variability in the dynamics of wound closure, averaging the percent area closure over time for 14 wounds of various shapes ranging from 0.02-0.125 mm2 allows the establishment of an average curve for wound healing in untreated animals (Figure 4E).
Damage to the basement membrane can be clearly seen when it occurs (Figure 2). In Movie 4, cells at the margin of a small wound in which there is basement membrane damage spread around the damaged area, and gap closure is completed with a purse string contraction.
If the tissue is dehydrated or too damaged to repair, cell movements can stop, or the entire sheet of cells can burst (Movie 5 and Movie 6). This usually happens after long periods of imaging (45 minutes or longer). If cell bursting occurs early in imaging, the sample is discarded.
As shown in Movie 7, large wounds heal in several stages. First, the edge of the wound becomes smooth and regular due to contractions at the margin, as previously reported57. Then, lamellipodia are seen to form from the cells at the wound margin, with lamellipodia moving forward to maximize contact with adjacent lamellipodia. Tracking of the nuclei in cells at the wound margin and several tiers behind the marginal cells shows that large gaps close by collective cell migration40. Cells never detach but move together as a sheet.
The introduction of dyes and pharmacological agents can be a powerful tool for dissecting biological mechanisms. Many substances are excluded from Clytia (not shown), likely because of the mucus layer that coats the surface of the animal. However, microinjection can be used to directly introduce molecules into the ECM, disrupting ECM structure or perturbing regulatory activities in the ECM. In addition, dyes and other molecules are able to enter epithelial cells from the basal side. For example, Figure 6 shows nuclear staining with Hoechst, membrane staining with FM1-43, and inhibition of lamellipodia formation by cytochalasin B after these reagents are microinjected into the ECM. The introduction of these molecules to the ECM and epithelial cells before wounding allows experiments that test the effect of pharmacological tools on the healing process.
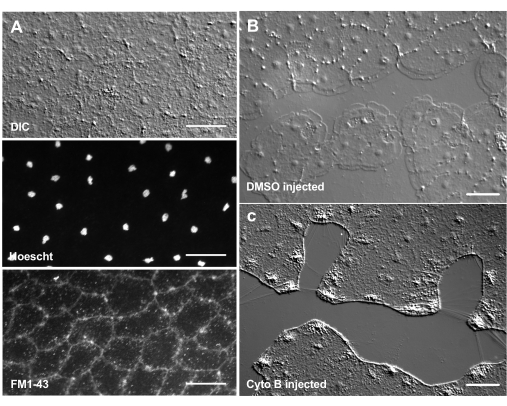
Figure 6: Epithelial cells of the medusa after microinjection of dyes or pharmacological agents. (A) Epithelial cells shown in top panel 5 min after injection with 20 µM Hoechst (nuclei) and 50 µM FM1-43 (membranes).(B,C) Wound healing after injection with 1:1,000 DMSO control (B) or 100 µM Cytochalasin B (C). Wounds were made 15 min after injection. Images were taken 5 min post-wounding. The formation of lamellipodia is inhibited by cytochalasin B. The apparent "fibers" often seen between cells in the wound area are believed to be the result of tension stretching the basement membrane — they do not stain with phalloidin (not shown). Scale bars: 50 µm. Please click here to view a larger version of this figure.
Movie 1: Time-lapse movie of single cell microwound healing. Time elapsed: 20 s. Frame rate: 10 fps. Scale bar: 50 µm. Please click here to download this Movie.
Movie 2: Time-lapse movie of a small epithelial wound healing. Time elapsed: 9 min 54 s. Frame rate: 10 fps. Scale bar: 50 µm. Please click here to download this Movie.
Movie 3: Time-lapse movie of a small epithelial wound healing. This wound is larger and more irregularly shaped than the wound in Movie 2. Time elapsed: 20 min 54 s. Frame rate: 10 fps. Scale bar: 50 µm. Please click here to download this Movie.
Movie 4: Time-lapse movie of a small wound and a microwound healing with a basement membrane tear. Lamellipodia spread around the basement membrane tear, although they can advance over the rest of the ECM. Once the region of the wound with the basement membrane damage is surrounded, a purse string contraction pulls cells over the region. Time elapsed: 19 min 4 s. Frame rate: 10 fps. Scale bar: 50 µm. Please click here to download this Movie.
Movie 5: Cells dying in a small epithelial wound. Cell death is likely due to dehydration of the animal. Time elapsed: 4 min 24 s. Frame rate: 10 fps. Scale bar: 100 µm. Please click here to download this Movie.
Movie 6: A small epithelial wound fails to complete healing. Time elapsed: 42 min 32 s. Frame rate: 10 fps. Scale bar: 50 µm. Please click here to download this Movie.
Movie 7: Large epithelial wound healing. Time elapsed: 25 min 29 s. Frame rate: 10 fps. Scale bar: 100 µm. Please click here to download this Movie.
Supplemental Figure 1: Clytia tank dimension schematics. 3D visualization of the custom-made Clytia tanks. (A) Front and back view. (B) Side view. The cut-out in the piece shown in green is covered with nylon mesh. Water enters the tank directly over the mesh, sweeps over the mesh and creates a circular current. Water exits the system through the hole in the end piece shown in blue.Please click here to download this File.
Supplemental Movie 1: Acellular extracellular matrix in Clytia. Z-stack of Clytia taken using confocal microscopy. The stack initially focuses on the exumbrella and then scans every 10 µm through the ECM to the plate endoderm and subumbrella. Images using DIC (left) and Hoechst nuclear staining (right) demonstrate the lack of cells in the ECM. Scale bar: 100 µm. Please click here to download this File.
