Nuclei were collected from fresh (unfixed) snap-frozen temporal neocortex tissue with a postmortem collection time of 12 h. After tissue dissociation into nuclei suspension, samples were incubated with antibodies against NeuN, PAX6, and OLIG2, and sorted according to the gating shown in Figure 1 and Figure 2. Nuclei were collected from NeuN+, PAX6+NeuN-, and OLIG2+NeuN- sorted populations (Figure 1E,F and Figure 2F). A targeted qPCR panel revealed enrichment for the pan-astrocyte markers, GFAP and ALDH1L1, in the PAX6+(NeuN-) population (Figure 1G and Figure 2H). Additionally, the OLIG2+ population was enriched for the oligodendrocyte progenitor cell (OPC) marker PDGFRA (Figure 1G). Bulk RNA sequencing of the collected populations showed comparative enrichment of astrocyte markers and depletion of neuronal markers in the PAX6+(NeuN-) population (Figure 1H). Immunofluorescence confirmed the colocalization of PAX6 with GFAP in adult human cortical astrocytes (Figure 1I).
Examining single-color controls reveals distinct positive and negative populations for each marker, which enabled the setting of accurate cutoffs for collection (Figure 2A–G). Furthermore, astrocyte-enriched FANS isolation was compared with two different antibodies against astrocyte nuclear markers, SOX9 and PAX6. A higher percentage of nuclei events was captured by FANS using PAX6 (~13.8%) than by FANS using SOX9 (~6%) within the astrocyte-enriched gate (Figure 2F,G). A targeted qPCR panel revealed enrichment for GFAP, PAX6, and SOX9 in both SOX9+(NeuN-) and PAX6+(NeuN-) populations (Figure 2H). Retroactively comparing the dissociation and staining of several samples with varying postmortem interval (PMI) of tissue collection revealed that a shorter PMI was associated with greater intact nuclei recovery (Figure 3A). Frozen tissue with a PMI of up to 24 h yielded a high rate of intact nuclei, up to 90%; at 30 h PMI, however, very few intact nuclei could be recovered (Figure 3A). Including an antibody-wash step in the standard protocol (which typically omits this step to optimize recovery) did not reveal significant shifts in the separation of distinct NeuN+/- or PAX6+/- populations (Figure 3B).
Occasionally, poor separation of PAX6+NeuN- populations could be seen, even in samples with a high percentage of viable nuclei (Figure 3C), necessitating the repetition of the tissue dissociation and the FANS protocol. Single-nucleus RNA-seq studies further prioritized PAX6 as a top differentially expressed nuclear transcription factor across both protoplasmic and fibrous adult astrocyte subpopulations, not only in the neocortex, but also in the subventricular zone (SVZ) and adjacent striatum (Figure 3D). Comparison of NEUN/PAX6/OLIG2 triple FANS between the neocortex and striatum derived from the same brain sample showed region-specific differences in the separation of PAX6+ nuclei (Figure 3E). This protocol is currently validated for the neocortex only.
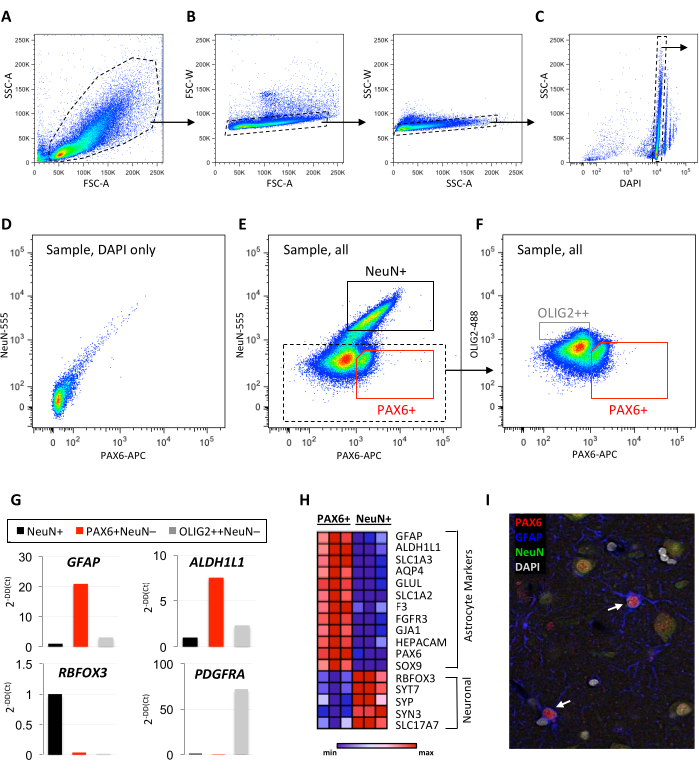
Figure 1: Validation of neuron- and astrocyte-enriched isolation by FANS. (A–F) Representative examples of sequential FANS gating to exclude debris and doublets (A–C) and define background staining using (D) a DAPI-only control for the collection of (E–F) enriched neuronal (NeuN+), astrocyte (PAX6+NeuN-), and OPC (OLIG2++NeuN-) nuclei populations. (G) Minimal qPCR panel using pan-astrocyte (GFAP, ALDH1L1), neuronal (RBFOX3), and OPC (PDGFRA) markers for quality control of the collected populations (A–G: temporal neocortex autopsy specimen without pathology, 12 h PMI, 100,000 events shown; dotted lines represent positively gated sequential populations; solid lines represent final collection of cell-type enriched populations). (H) Heatmap generated from row-normalized bulk RNA sequencing data showing expression of canonical astrocyte and neuronal markers in PAX6+ and NeuN+ sorted populations (n=3 autopsy cases, temporal neocortex without pathology, PMI 12-21 h). (I) Representative immunofluorescence image of PAX6, GFAP, and NeuN in human neocortex. Arrows indicate astrocytes co-expressing PAX6 and GFAP. Abbreviations: FANS = fluorescence-activate nuclei sorting; DAPI = 4′,6-diamidino-2-phenylindole; NeuN = neuronal nuclei; PAX6 = paired box protein 6; OLIG2 = oligodendrocyte transcription factor 2; OPC = oligodendrocyte progenitor cell; qPCR = quantitative polymerase chain reaction; GFAP = glial fibrillary acidic protein ; ALDH1L1 = 10-formyltetrahydrofolate dehydrogenase; RBFOX3 = RNA-binding protein FOX-1 homolog 3; PDGFRA = platelet-derived growth factor alpha; PMI = postmortem interval; SSC-A = side scatter-area; FSC-A = forward scatter area; SSC-W = side scatter width; FSC-W = forward scatter width; NeuN-555 = mouse anti-NeuN conjugated to AF555; PAX6-APC = mouse anti-PAX6 conjugated to allophycocyanin; OLIG2-488 = mouse anti-OLIG2 conjugated to green fluorescent dye for the 488 nm laser line. Please click here to view a larger version of this figure.
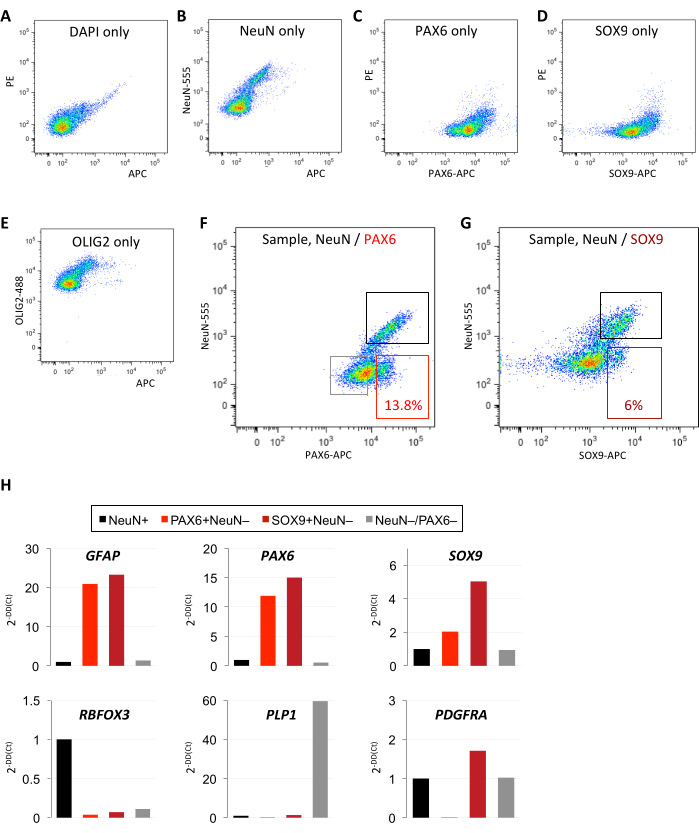
Figure 2: Comparative isolation of astrocyte-enriched populations using PAX6 or SOX9 antibodies. (A–E) Single-color controls for DAPI, NeuN, PAX6, SOX9, and OLIG2 define positive/negative cutoffs in the respective fluorescence channels. (F–G) Comparative FANS method for astrocyte-enriched isolation using (F) NeuN/PAX6 or (G) NeuN/SOX9 antibodies. A greater percentage of events are captured by PAX6 than by SOX9 within the astrocyte-enriched gate. (A–G: identical postmortem non-pathological temporal neocortex specimen used for all experiments; 12 h PMI; 10,000 events shown). (H) Quality control qPCR analysis confirms enrichment of astrocyte markers and depletion of non-astrocyte markers in PAX6+(NeuN-) and SOX9+(NeuN-) sorted populations from F–G (data normalized to ACTB and quantified as fold change of the NeuN+ population). Abbreviations: DAPI = 4′,6-diamidino-2-phenylindole; NeuN = neuronal nuclei; PAX6 = paired box protein 6; OLIG2 = oligodendrocyte transcription factor 2; SOX9 = SRY-box transcription factor 9; qPCR = quantitative polymerase chain reaction; GFAP = glial fibrillary acidic protein ; RBFOX3 = RNA-binding protein FOX-1 homolog 3; PDGFRA = platelet-derived growth factor alpha; PMI = postmortem interval; NeuN-555 = mouse anti-NeuN conjugated to AF555; PAX6-APC = mouse anti-PAX6 conjugated to allophycocyanin; OLIG2-488 = mouse anti-OLIG2 conjugated to green fluorescent dye for the 488 nm laser line. Please click here to view a larger version of this figure.
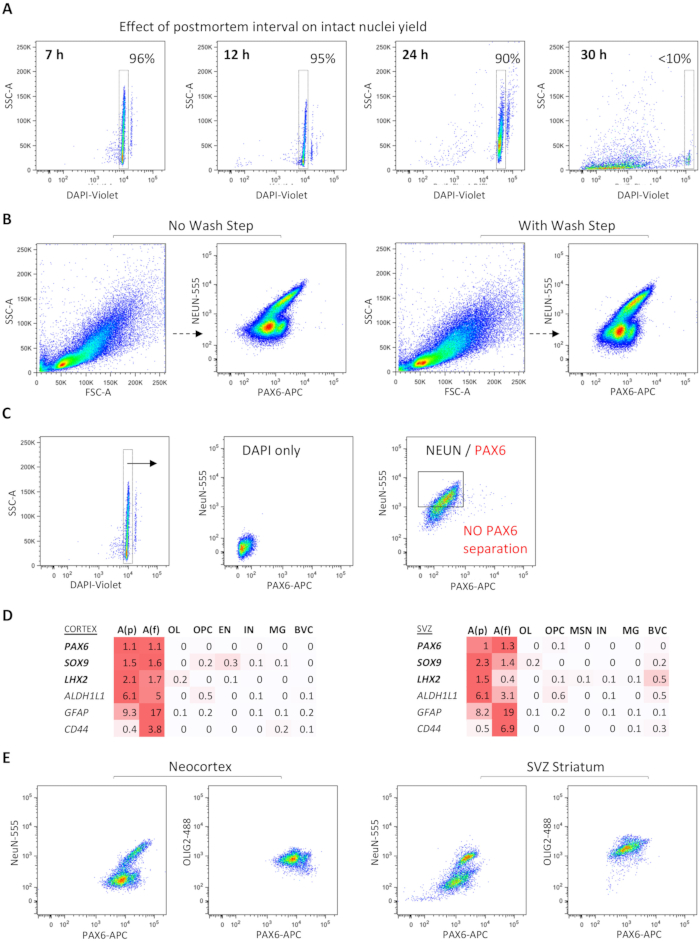
Figure 3: Dependent and independent metrics for successful FANS experiments. (A) Effect of sample collection PMI on the quality of nuclei: lower PMI enables the recovery of a greater number of intact nuclei with adequate results up to 24 h. (B) The inclusion of a wash step after antibody incubation does not visually appear to shift FANS populations. (C) Example of a failed FANS experiment with lack of distinct PAX6+(NeuN-) population (on right) despite the presence of intact nuclei and low background (on left) (postmortem temporal neocortex, non-pathological, 7 h PMI). (D) Heatmap representation of adult human sn RNA-seq data showing differential expression of PAX6 and two other astrocyte nuclear factors, SOX9 and LHX2, across protoplasmic and fibrous astrocyte subpopulations in both cortex and SVZ, compared to other cell types (red color gradient represents spectrum of log-normalized average gene expression values; n=3 distinct autopsy cases, 43,619 total nuclei. (E) Distinct brain regions (temporal neocortex vs. SVZ and subjacent striatum) show different patterns of NeuN/PAX6/OLIG2 separation. Abbreviations: FANS = fluorescence-activate nuclei sorting; DAPI = 4′,6-diamidino-2-phenylindole; NeuN = neuronal nuclei; PAX6 = paired box protein 6; OLIG2 = oligodendrocyte transcription factor 2; GFAP = glial fibrillary acidic protein ; ALDH1L1 = 10-formyltetrahydrofolate dehydrogenase; LXH2 = LIM homeobox 2; PMI = postmortem interval; SSC-A = side scatter-area; FSC-A = forward scatter area; NeuN-555 = mouse anti-NeuN conjugated to AF555; PAX6-APC = mouse anti-PAX6 conjugated to allophycocyanin; OLIG2-488 = mouse anti-OLIG2 conjugated to green fluorescent dye for the 488 nm laser line; SVZ = subventricular zone; A(p) = protoplasmic (gray matter) astrocytes; A(f) = fibrous (white matter) astrocytes; OL = oligodendrocytes; OPC = oligodendrocyte progenitor cells; EN = excitatory neurons; MSN = medium spiny neurons; MG = microglia; BVC = blood vessel cells. Please click here to view a larger version of this figure.
