Mice have one CVP, located posteriorly on the tongue, from which LGR5+ stem cells can be isolated (Figure 1A, black box). Injection of an enzyme solution under and around the CVP (Figure 1B) results in slight swelling of the epithelium and digestion of the connective tissue. Sufficient digestion is achieved following a 33 min incubation, which allows easy separation of the CVP epithelium from the underlying tissue. When attempting to peel the CVP epithelium, cuts should be made at a sufficient distance from the CVP to ensure trenches are not disrupted or damaged (Figure 1C). This also enables one to grip the epithelium using forceps without damaging the CVP. Trimming the epithelium surrounding the CVP removes non-target cells and increases the efficiency of the following steps by decreasing the tissue mass being manipulated (Figure 1D). It is important to check that the two trenches (Figure 1C, black arrows) are present before adding CVP epithelium to the microcentrifuge tube; successful peeling of the CVP includes part of the Von Ebner's glands and ducts (Figure 1D, black arrows). If the peeled epithelium does not contain these two opaque structures, the trench epithelium is most likely ruptured due to incomplete digestion of the connective tissue.
To establish proper FACS settings to collect Lgr5-EGFP cells, cells from dissociated CVPs are separately obtained from wild type and Lgr5-EGFP mice. Wild type CVP cells are analyzed first to establish gating parameters, a process in which populations of cells are categorized within the scatterplot output by characteristics of interest24. Here, four parameters were used to ultimately identify cells for plating. The first parameter, forward scatter, filters out particles and debris based on the detected surface area. This parameter removes ~70% of all detected events during the sort (Figure 3). The width parameter further filters events based on size to ensure selection of single cells (singlets). Approximately 90% of events are singlets (Figure 3). The nuclear marker DAPI is taken up by dead but not live cells and thus allows dead cells to be sorted out25. This protocol optimizes cell viability, as over 90% of events are live cells (Figure 3). Lastly, GFP gating parameters are set above the autofluorescence level of wild type cells. Wild type cells are not collected; they are used solely as a gating control for GFP fluorescence. With gating parameters determined from the wild type sample, cells from the Lgr5–EGFP sample are then run through the flow cytometer to be sorted for collection. Gates can be adjusted at the beginning of the Lgr5-EGFP sort to accommodate clear clustering of certain cell populations but should not be significantly changed mid-sort. It was found that the dissociation of three pooled Lgr5-EGFP CVPs yields ~500,000 cells, including an average of 10,000 GFP+ cells.
Proper media and culture conditions are vital for optimal growth and differentiation of organoids. Previous studies utilizing lingual organoids modeled their media components after those from the intestinal organoid field, including Wnt3a, EGF, Noggin, and R-spondin17,18,19,20,21,22. However, when lingual organoids are cultured using a similar method (WENR media), organoids do not grow efficiently (Figure 5A). In human intestinal organoids, drugs A8301 (a TGFß signaling inhibitor) and SB202190 (a p38 MAPKinase signaling inhibitor) are used to promote organoid growth26. Indeed, adding these inhibitors (WENRAS media) induces robust growth of lingual organoids (Figure 5A). Interestingly, removing these inhibitors from the media after 6 days results in higher expression of general TRC marker Kcnq1, suggesting A8301 and SB202190 hinder taste cell differentiation (Figure 5B). Thus, optimal growth and differentiation are obtained by culturing organoids in WENRAS media from days 0-6 and WENR media from days 6-12 (Figure 4, Figure 5), respectively.
Organoid cell type composition can be characterized by qRT-PCR or immunohistochemistry. Mature organoids contain both taste cells, marked by Kcnq1 and KRT8, and non-taste epithelial cells, marked by Krt13/KRT13 (Figure 6, 8A). This suggests isolated LGR5+ cells have a similar potency in vitro as they do in vivo since, in the adult tongue, Lgr5-GFP+ cells produce both taste and non-taste lineages5. Further, Krt13 is expressed at higher levels than all 3 TRC markers (Figure 6), suggesting organoids are predominately composed of non-taste epithelial cells. In fact, relative quantification of gene expression27 indicates Krt13 is expressed 50x higher than general TRC marker Kcnq1 (Student's t-test, p = 0.0004). This is expected, as the tongue has a similar proportion of taste versus non-taste epithelium1. Organoids express all three TRC types (Figure 6, 8B,C). Type I cells (marked by Entpd2) and bitter type II cells (marked by Gnat3) are highly expressed in taste organoids, while sour sensing type III cells (marked by Car4) are less common (Figure 6). TRCs are randomly distributed in organoids (Figure 8) rather than in discrete taste bud structures observed in vivo.

Figure 1: Dissected tongue and peeled CVP epithelium. (A) The tongue is dissected out, and the anterior tongue is removed by cutting just anterior of the intermolar eminence (dashed line), leaving the posterior tongue, which includes the CVP (black box) (B) A needle is inserted just anterior to the intermolar eminence (black arrow), and enzyme mixture is injected below and to the lateral edges of the CVP (black box). (C) Untrimmed peeled epithelium surrounding the CVP trenches (black arrows) and (D) trimmed peeled CVP epithelium. Von Ebner's glands and ducts (D, black arrows) are visible after successful peeling of the trenches. Scale bar: 1 mm. Please click here to view a larger version of this figure.
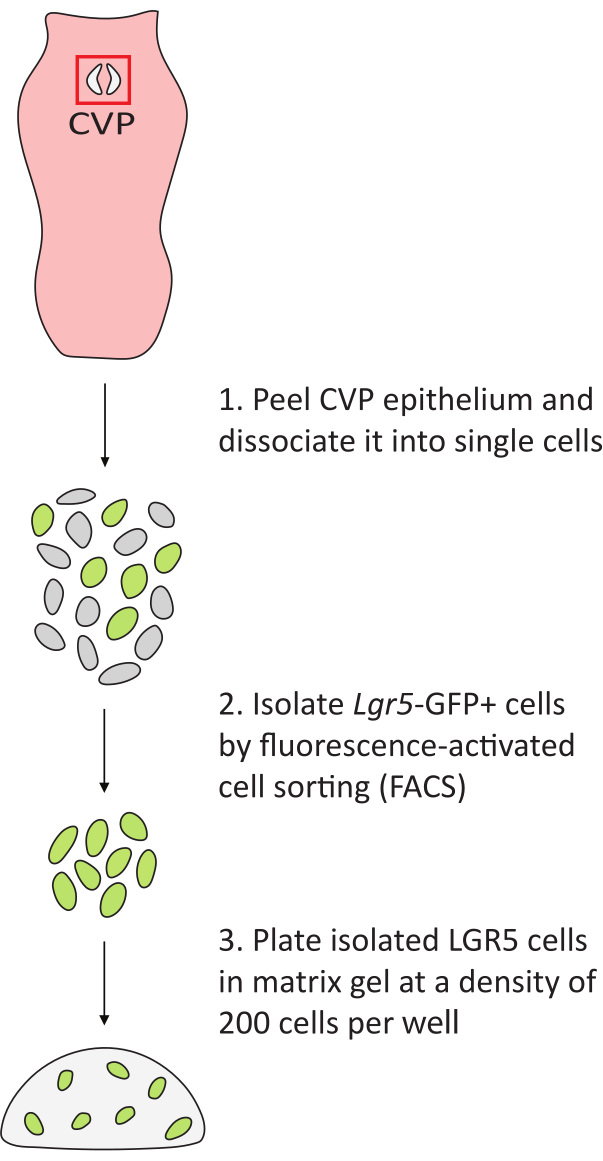
Figure 2: Workflow of lingual organoid generation. The tongue is removed from Lgr5–EGFP mice. The CVP trench epithelia (red box) are peeled from the underlying connective tissue and dissociated into single cells. GFP+ cells are isolated and plated in matrix gel at a density of 200 cells per well in a 48-well plate. Please click here to view a larger version of this figure.
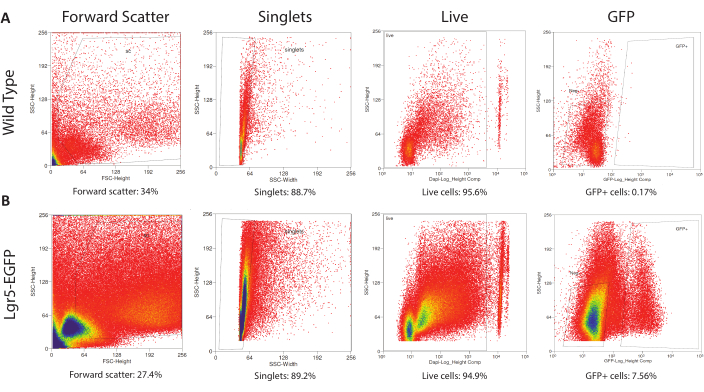
Figure 3: Gating for Fluorescence-Activated Cell Sorting. (A) The control (wild type CVP cells) run determines the FACS gates that eliminate debris and broken cells via forward scatter, identifies singlets via side scatter width, separates DAPIneg live from DAPI+ dead cells, and establishes the autofluorescence level of wild type cells. (B) Previously determined gates are applied to the experimental run (Lgr5EGFP-IRES-CreERT2 CVP cells) to isolate debris-free, single, live, Lgr5-EGFP+ cells. Please click here to view a larger version of this figure.
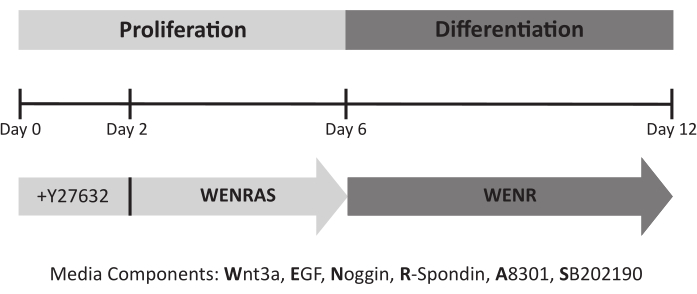
Figure 4: Lingual organoid culture timeline and required media. ROCK inhibitor Y27632 is added to the media for the first 48 h of culture to promote cell survival. During the proliferation phase, organoids are fed conditioned medium containing A8301 and SB202190 (WENRAS) to optimize growth. These drugs are withheld from media (WENR) starting at day 6 to promote differentiation. Please click here to view a larger version of this figure.
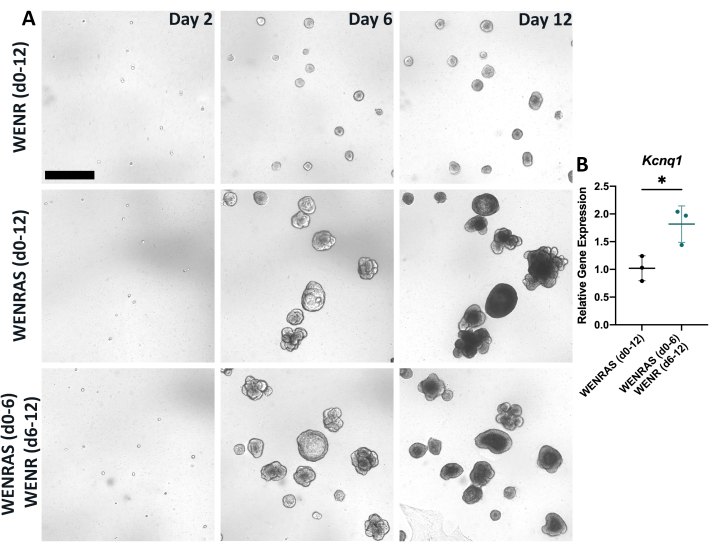
Figure 5: Drugs A8301 and SB202190 affect organoid growth and differentiation. (A) Brightfield images of organoids grown in either WENR media (day 0-12), WENRAS media (day 0-12), or WENRAS (day 0-6), then WENR (day 6-12). Images were captured at day 2, day 6, and day 12 of culture using live imaging software. Scale bar: 400 µm (B) Relative gene expression of a global TRC marker Kcnq1 is significantly reduced when drugs A8301 and SB202190 are present during organoid differentiation. Each point represents one biological replicate, which included three pooled wells. Mean change in relative gene expression (horizontal line) was calculated by averaging three biological replicates. Error bars: ±SD. Please click here to view a larger version of this figure.
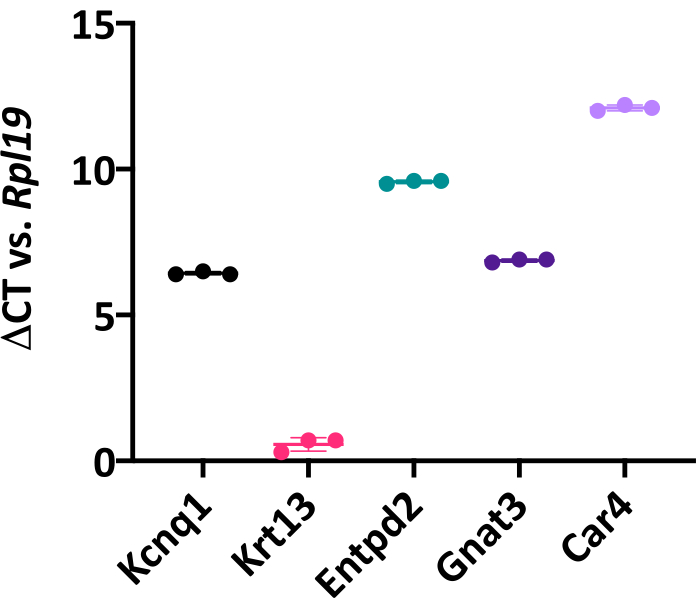
Figure 6: Lingual organoids express canonical taste cell markers. Change in cycle threshold (Ct) value of global TRC marker Kcnq1, non-taste epithelial marker cytokeratin 13 (Krt13), type I TRC marker Entpd2, type II bitter TRC marker Gnat3, and type III sour TRC marker Car4, compared to housekeeping gene Rpl19. Each point represents one biological replicate, which included three pooled wells from a 48-well plate. Mean change in Ct value (horizontal line) for each marker was calculated by averaging three biological replicates. Unpaired Student's t-test, *p < 0.05. Error bars: ± SD. Please click here to view a larger version of this figure.

Figure 7: Mounting of lingual organoids for inverted confocal microscopy. (A) An ~1 mm thick string of non-toxic modeling clay is created. (B) The clay is sculpted as an ~20 mm x 20 mm square in the center of a 24 mm x 75 mm microscope slide. (C) A 22 mm x 22 mm square coverslip seals organoids suspended in mounting medium. Scale bar: 10 mm. Please click here to view a larger version of this figure.
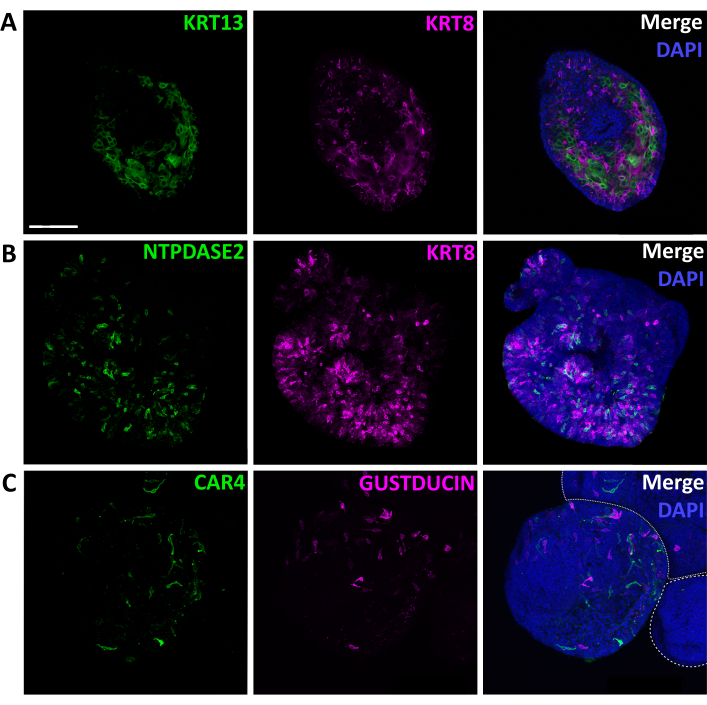
Figure 8: Immunolabeling of intact lingual organoids. Confocal images of fixed, immunostained organoids. (A) Optical section of an organoid stained for non-taste epithelial marker KRT13 (green) and general TRC marker KRT8 (magenta). (B) Maximum projection of a confocal z-stack of one organoid stained for type I glial-like cell marker NTPDase2 (green) and KRT8 (magenta). (C) Maximum projection of a confocal z-stack of two partially shown organoids (white-dashed outlines) and one complete organoid stained for type III sour detecting cell marker CAR4 (green), and bitter detecting type II cell marker GUSTDUCIN (magenta). Scale bar: 100 µm for A, B, and C. Nuclear marker DAPI (blue, right column). Please click here to view a larger version of this figure.
| Gene Name | Forward primer (5'-3') | Reverse primer (5'-3') | Assession Number | ||||||
| Car4 | CTGCTAGGACAAAGGTGAACC | CTCCACTGTGTGTTGATTGTTCT | NM_007607.2 | ||||||
| Entpd2 | GACAAGGAAAATGACACAGGTATCGTGG | GTTCAAGACATTCAACCAGACTC | NM_009849.2 | ||||||
| Gnat3 | ATCCAGGAATCCAAGCCTGC | TGGTTTTCACCCGGGAATGT | NM_001081143.1 | ||||||
| Kcnq1 | TTTGTTCATCCCCATCTCAG | GTTGCTGGGTAGGAAGAG | NM_008434.2 | ||||||
| Krt13 | TCATCTCGGTTTGTCACTGGA | TGATCTTCTCGTTGCCAGAGAG | NM_010662.2 | ||||||
| Rpl19 | GGTCTGGTTGGATCCCAATG | CCCGGGAATGGACAGTCA | NM_009078.2 | ||||||
Table 1: Primer sequences used for quantitative RT-PCR.
Supplementary Table 1: Comparison of published lingual organoid culture media components. Please click here to download this Table.
