The goal of this study was to develop a simple protocol for directly introducing recombinant viruses engineered for gene silencing or gene expression into maize seedlings (Figure 2). The virus vectors carrying inserts are designed and cloned using standard molecular biology techniques. Gene fragments for silencing are inserted into MCS1* in FoMV-EV and coding sequences for expression are inserted into FoMV-EV at MCS2 or SCMV-EV at MCS. The resulting plasmids are transferred to Agrobacterium strain GV3101. Subsequently, maize seedlings are injected within a week or less after planting. Two weeks after injection, plants can be assessed both phenotypically and molecularly for viral infection, gene silencing, and gene expression.
Maize plants are grown in a peat-based medium for 4-7 days. At this stage, the shoot apical meristem is just above the coleoptilar node (Figure 3A). After the coleoptile has extended 2-3 centimeters or up until 7 days after sowing, plants are injected 2-3 mm above the coleoptilar node (Figure 3B-F). At approximately 12 days after injection, plants will begin displaying silencing phenotypes on their leaves, commonly observed near vascular tissue, and these lesions are visually distinct from FoMV viral mosaic symptoms (Figure 4). Both the presence of FoMV and the silencing of target genes is detectable in injected plants (Figure 5). GFP expression can be detected by 2 weeks after injection under a fluorescent microscope and is strongest on leaves 5-7 (Figure 6). When observed under a fluorescence imaging system, GFP expression from FoMV can be visualized as many small, punctate areas of fluorescence distributed across leaves near vascular tissue while GFP expression from SCMV consists of larger patches (Figure 6, Supplemental Figure 1). Although viral mosaic symptoms are often visible on plants infected with FoMV silencing constructs, plants injected with GFP expression constructs that are successfully expressing GFP often do not have these symptoms. As a result, a plant with no visible symptoms may still be positive for virus and GFP expression. Additionally, puncturing the meristem during the agroinjection procedure should be avoided as this can cause morphological defects, but the resulting plants survive and are often symptomatic (Figure 7).
Although this protocol was originally developed using sweet corn, several maize inbred lines can be successfully inoculated with FoMV gene silencing constructs using agroinjection. For example, FR1064 and B73 typically have high rates of viral infection (Table 2). Notably, Mo17, a line with known genetic resistance to FoMV, had a 0% infection efficiency as expected36, 53. Additionally, the construct used influences infection efficiency (Table 3). In the case of FoMV, FoMV-EV and FoMV-LES22 typically have the highest infection efficiencies at 53% and 54%, respectively. FoMV-PDS has a slightly lower efficiency at 38%, and FoMV-GFP is the lowest at 17%. SCMV-GFP has an infection efficiency of 8%. These percentages are averages over several experiments; individual experiments can have higher or lower infection efficiencies.
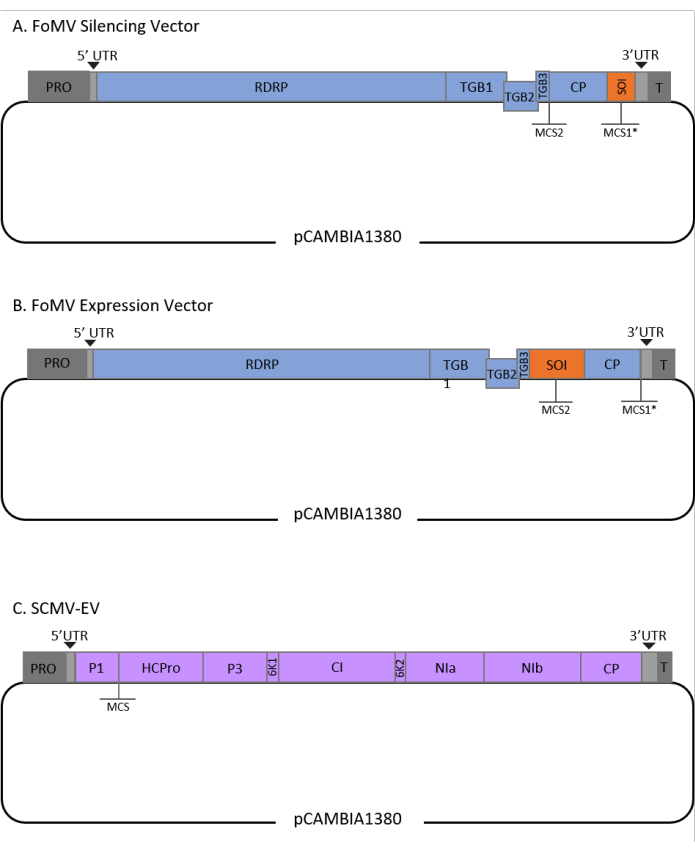
Figure 1: Schematic representations of the FoMV and SCMV T-DNA clones used for agroinjection in maize. The FoMV vector contains two multiple cloning sites (MCS1* and MCS2). The empty vector, FoMV-EV, is 7,269 bp and does not contain any inserts in either MCS. (A) Gene silencing using the FoMV vector can be achieved by inserting gene fragments into MCS1*, designated as sequence of interest (SOI), typically in the anti-sense orientation. (B) Gene expression using the FoMV vector can be accomplished by inserting gene ORFs into the MCS2 in the sense orientation, designated as SOI. (C) The SCMV vector was engineered to have one MCS between P1 and HCPro. The empty vector, SCMV-EV, is 11,015 bp and does not contain any inserts in the MCS. Gene ORFs inserted into the MCS that are in frame with the SCMV polyprotein will be expressed as proteins. Please click here to view a larger version of this figure.
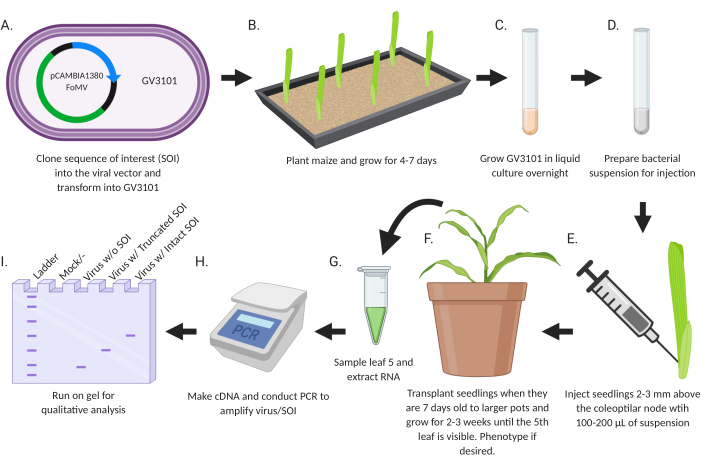
Figure 2: Schematic summary of the agroinjection protocol. (A) Clone SOI, either a CDS or gene fragment, into the viral vector and transform into Agrobacterium strain GV3101. (B) Plant maize and grow for 4-7 days. (C) Grow GV3101 in liquid culture overnight at 28 °C. (D) Prepare bacterial suspension for injection. (E) Inject seedlings 2-3 mm above the coleoptilar node with 100-200 µL of suspension. (F) Transplant seedlings when they are 7 days old to larger pots and grow for 2-3 weeks until the 5th leaf is visible. Phenotype if desired. (G) Sample leaf 5 and extract RNA. (H) Make cDNA and conduct PCR to amplify virus/SOI. (I) Run on gel for qualitative analysis to determine presence/absence of virus and a truncated or intact SOI. This figure was created with BioRender.com. Please click here to view a larger version of this figure.

Figure 3: Agroinjection method of inoculating seedlings just above the coleoptile node. (A) 4-5-day old plants. The coleoptile is fully expanded, and the first true leaf may be partially visible, but is not unfurled. (B) 6-7-day old plants. The first leaf may be expanded but no collars will be visible. The second leaf will also be visible and may be starting to unfurl at this stage. (C) Dissection of 6-7-day old plants showing the location of the shoot apical meristem in relation to the coleoptile node. (D) Injection of 4-5-day old plants. (E) Injection of 6-7-day old plants. (F) Injection of 6-7-day old plants using a dye solution, showing dyed inoculum coming out of the whorl of the seedling. (G) Close-up of injection site of 6-7-day old plants in relation to the coleoptile node. (H) Close-up of a 6-7-day old plant post-injection, showing dyed inoculum in the whorl of the plant. Please click here to view a larger version of this figure.

Figure 4: Symptoms of the silencing control genes used in the agroinjection experiments. (A) A leaf photographed at 17 DPI after the plant was injected with FoMV-LES22. FoMV-LES22 carries a 329 bp insert of the 3' CDS of the lesion mimic 22 maize gene in the antisense orientation. Silencing results in the accumulation of a toxic metabolite which in turn causes the necrotic lesions that first appear as streaks along vasculature and grow into larger patches as shown here. (B) A leaf photographed at 17 DPI after the plant was injected with FoMV-PDS. FoMV-PDS carries a 313 base pair insert of the 3' CDS of the sorghum phytoene desaturase gene in the antisense orientation. Silencing of pds in maize causes a systemic photobleaching phenotype that starts as small, thin streaks along vasculature that grow into longer streaks along the length of the leaf as shown here. Please click here to view a larger version of this figure.
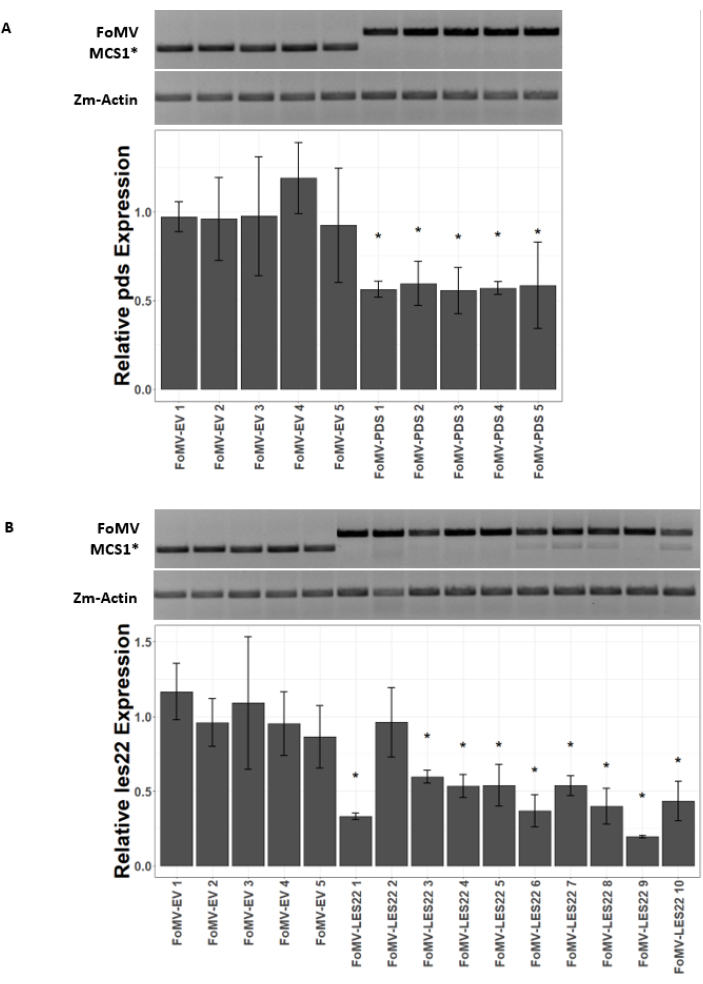
Figure 5: qRT-PCR of plants injected with FoMV gene silencing constructs. Confirmation of systemic FoMV infection and gene silencing induced by the FoMV-LES22 and FoMV-PDS constructs delivered via agroinjection in sweet corn plants (Golden x Bantam). (A) The gel images show RT-PCR analyses confirming the presence of FoMV-MCS1* empty vector (315 bp amplicon) and FoMV-PDS (625 bp amplicon) in leaf 6 of five individual plants. The PCR primers used produce an amplicon that spans MCS1*. The maize gene actin (Zm-Actin) amplicon serves as the reference gene. The bar graph represents the qRT-PCR relative expression values for pds expression in leaf 6 at 37 days post inoculation (DPI) by agroinjection with FoMV-MCS1* or FoMV-PDS. Suppression of pds is detectable in each of the five biological replicates (p=0.003; post hoc Dunnett's test; error bars indicate standard deviation (SD) of three technical replicates). (B) The gel images show RT-PCR analyses confirming the presence of FoMV-MCS1* (315 bp amplicon) in leaf 6 of five individual plants. FoMV-LES22 (625 bp amplicon) was detected in leaf 6 tissue (samples FoMV-LES22 1-5, 38 DPI) and leaf 4 (samples FoMV-LES22 6-10, 20 DPI) for ten individual plants. The Zm-Actin amplicon served as the reference gene. The bar graph represents the qRT-PCR relative expression values for les22 expression in maize tissues by agroinjection of FoMV-MCS1* or FoMV-LES22 viral constructs. Les22 suppression occurs in 9 of 10 biological replicates (p=<0.0001; post hoc Dunnett's test; error bars indicate SD for three technical replicates). Please click here to view a larger version of this figure.

Figure 6. Phenotypes of various constructs used in the agroinjection experiments. All imaged plants were injected when they were 6-7 days old with Agrobacterium strain GV3101 carrying the indicated constructs. Images were taken at 16 DPI. (A) Leaf symptoms of pCAMBIA1380 (empty plasmid backbone), FoMV-EV, FoMV-GFP, and SCMV-GFP in visible light, under the FluorCam chlorophyll filter at 250 µs exposure, and under the FluorCam GFP filter at 10 ms exposure. (B) Fluorescent microscopy images of the leaves of mock-treated (injected with MgSO4 solution only), FoMV-EV, and FoMV-GFP injected plants. The DIC, DsRed, and EGFP channels are shown and were each taken at 1500 ms exposure. Scale bar is 200 µm. Please click here to view a larger version of this figure.

Figure 7. Morphological effects of injection. An example of the more severe morphological effects that can occur from direct injection into meristematic tissue. This injury can result in "shredding" of the leaves and splitting of the stem. Please click here to view a larger version of this figure.
| Virus | Growth Conditions | Genotype | # Infected Plants | Total # of Plants | % Infection | Avg % of Infection |
| FoMV-EV | Growth Chamber | Sweet Corn | 22 | 23 | 96% | 97% |
| B73 | 18 | 18 | 100% | |||
| B104 | 20 | 21 | 95% | |||
| Greenhouse | Sweet Corn | 20 | 23 | 87% | 89% | |
| B73 | 17 | 18 | 94% | |||
| B104 | 16 | 19 | 84% | |||
| SCMV-EV | Growth Chamber | Sweet Corn | 14 | 21 | 67% | 47% |
| B73 | 5 | 18 | 28% | |||
| B104 | 10 | 21 | 48% | |||
| Greenhouse | Sweet Corn | 14 | 23 | 61% | 49% | |
| B73 | 0 | 19 | 0% | |||
| B104 | 19 | 22 | 86% |
Table 1: Effect of greenhouse and growth chamber conditions on agroinjection inoculation efficiency. Seeds were germinated under identical growth conditions. Germinated seedlings were agroinjected and half of them were moved to a growth chamber (25 °C 16 h daylight/ 22C 8 h night; 185 PAR) and the other half were moved to a greenhouse (22-25 °C 16 h daylight/22-25 °C 8 h night; 350-400 PAR). This table reports the rate of infection as a percentage, calculated from the number of plants confirmed by RT-PCR to be infected with the respective virus divided by the total number of agroinjected plants. There is no statistical difference in infection efficiencies between growth chamber and greenhouse conditions (FoMV two tailed t-test p=0.08; SCMV two tailed t-test p=0.96).
| Maize Genotype | FoMV-EV | FoMV-LES22 | Combined Total | ||||
| Infected | Total | % Infected | Infected | Total | % Infected | % Infected | |
| Sweet Corn | 18 | 23 | 78% | 15 | 23 | 65% | 72% |
| MO47 | 7 | 22 | 32% | 1 | 21 | 5% | 19% |
| K55 | 1 | 15 | 7% | 3 | 17 | 18% | 13% |
| W64A | 10 | 22 | 45% | 8 | 20 | 40% | 43% |
| MO17 | 0 | 16 | 0% | 0 | 13 | 0% | 0% |
| B73 | 10 | 18 | 56% | 7 | 17 | 41% | 49% |
| B101 | 12 | 21 | 57% | 8 | 24 | 33% | 44% |
| FR1064 | 4 | 4 | 100% | 4 | 4 | 100% | 100% |
| B104 | 10 | 22 | 45% | 5 | 21 | 24% | 35% |
| WCC22 | 2 | 7 | 29% | 4 | 6 | 67% | 46% |
| A188 | 0 | 3 | 0% | 4 | 6 | 67% | 44% |
Table 2: Infection efficiency of FoMV constructs across maize genotypes. FoMV-EV and FoMV-LES22 were agroinjected into 11 genotypes of maize. After injection, the seedlings were moved to the greenhouse. This table details the rate of infection as a percent, calculated from the number of plants infected with FoMV as confirmed by RT-PCR divided by the total number of agroinjected plants. The combined total rate of infection shows the average rates of infection of each genotype for both FoMV constructs tested.
| Plant Stage | 4-5 Day Old Plants | 6-7 Day Old Plants | Combined Total | ||||
| Symptomatic | Total Plants | % Infected | Symptomatic | Total Plants | % Infected | % Infected | |
| FoMV-EV | 42 | 72 | 58% | 80 | 170 | 47% | 53% (A) |
| FoMV-PDS | 65 | 157 | 41% | 66 | 184 | 36% | 39% (B C) |
| FoMV-LES22 | 115 | 195 | 59% | 144 | 292 | 49% | 54% (A B) |
| FoMV-GFP | 16 | 103 | 16% | 37 | 217 | 17% | 16% (C) |
| SCMV-GFP | 10 | 95 | 11% | 5 | 82 | 6% | 8% (C) |
Table 3: Summary of injection experiments. This table represents a summary of the injection experiments conducted from August 2017 to August 2018 on Golden Bantam sweet corn seedlings. Plants were assessed for viral symptoms (FoMV-EV), silencing symptoms (pds and les22) or GFP fluorescence (GFP) through visual (FoMV-EV, FoMV-PDS, and FoMV-LES22) or FluorCam (FoMV-GFP and SCMV-GFP) screening. Results are shown individually for 4-5 day old plants and 6-7 day old plants, as well as a summary across all plant ages. There is no significant difference found between 4-5 day old plants and 6-7 day old plants (One-way ANOVA, F=0.6513). There is a difference found between viral construct (Onaway ANOVA, F=<0.0001), with the letters representing the Tukey-Kramer HSD connecting letters report.
Supplemental Table 1: Table listing all primer names and sequences used in this protocol. Please click here to download this Table.
Supplemental Table 2: Acetosyringone test. (A) Initial acetosyringone test, comparing rates of symptoms of mock, FoMV-EV, and FoMV-LES22 injected plants between inoculation suspensions with 200 µM acetosyringone (+) or without acetosyringone (-). (B) Comparing the rates of infection of FoMV-LES22 as determined by RT-PCR between inoculation suspensions without acetosyringone (-), with 200 µM acetosyringone (+), and addition of 20 µM of acetosyringone to the bacterial culture 4 hours prior to resuspension in buffer along with the addition of 200 uM acetosyringone to the final suspension (++). Overall, there was no significant difference found between aceotysyringone treatments (Oneway ANOVA, f=0.5452). Please click here to download this Table.
Supplemental Figure 1: Fluorescence imaging and molecular validation of agroinjected SCMV and expression of heterologous proteins in maize. Maize was agroinjected with a modified SCMV construct containing both CDSs of GFP and nano luciferase (NLuc). (A) Fluorcam imaging was used for screening and detection of GFP. The left is a mock injected plant and the right is SCMV-NLucGFP injected plant. (B) Leaf protein extracts were separated by SDS-PAGE and evaluated for the presence of NLuc, GFP, and SCMV coat protein (CP) by in-gel luciferase assay or immunoblot as indicated. Please click here to download this File.
