All mouse procedures were approved by the Institutional Animal Care and Use Committee of Washington University. Mice of any sex, age, and weight may be used for these experiments and will be dependent on the nature of the experimental question one seeks to address. Mice used here are adult (12-16 weeks old), male wild-type C57BL/6 mice. All human procedures were approved by the Institutional Review Board of Washington University. Study subjects consented to data usage, and representative human subject data included within this protocol are from a published study14. Data here is from non-diabetic post-menopausal (55-75 years old) females. Details for preparing reagents necessary for the assay are presented in Table 1. Information on the specific reagents, tools, and machines used in the assay are listed in the Table of Materials. A schematic overview of the protocol is presented in Figure 1.
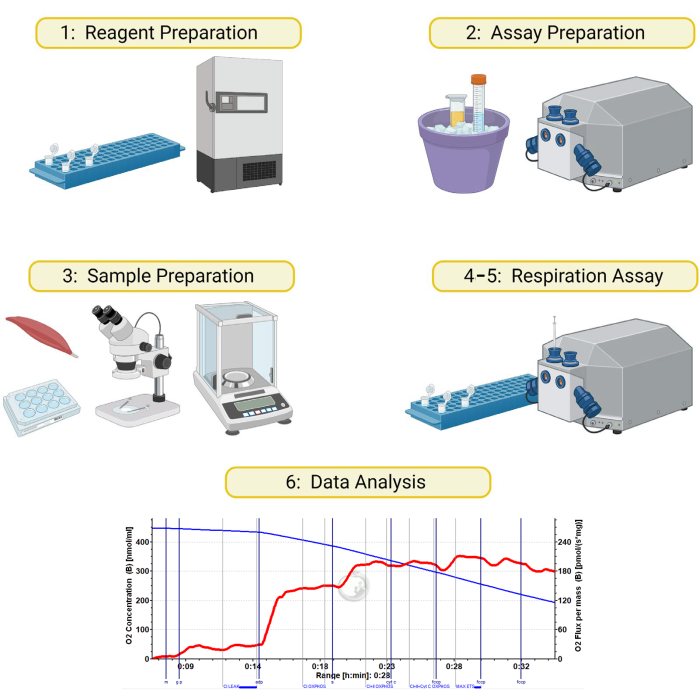
Figure 1: Schematic for high-resolution respirometry on permeabilized skeletal muscle samples. The method detailed in this manuscript is divided into 6 sections: 1) preparation of respiration buffers and reagents, 2) instrument and reagent preparation on day of assay, 3) preparation and permeabilization of muscle samples, 4) preparing sample and instrument, 5) running the respiration assay, and 6) data analysis. Created with BioRender.com Please click here to view a larger version of this figure.
1. Assay preparation and instrument calibration
- On the day of the assay, remove the needed number of aliquots of each respiration compound (blebbistatin, palmitoyl-carnitine, glutamate, malate, ADP, succinate, cytochrome C, FCCP) and thaw on ice. Thaw vials of BIOPS and MiR05 on ice.
- Prepare saponin and pyruvate solutions as indicated in Table 1. Prepare a working solution of MiR05 as in Table 1. Prepare 5 mL of working solution of MiR05 per instrument (2 chambers). Scale up as needed.
- Turn on the high-resolution respirometry system and the vacuum system. Remove the stoppers, remove the 70% ethanol storage solution by vacuum, and refill the chamber with ultrapure molecular grade H2O. Remove H2O by vacuum and refill. Repeat for a total of 3 washes. After the final wash, add 2 mL of MiR05 (without creatine or blebbistatin) to each chamber.
- Open the respirometry software. After the software initiates, a pop-up box will open for instrument settings. Set chamber stir speed to 750 rpm, temperature to 37 °C, and data recording interval to 2 s. Set gain to 1 and polarization voltage to 800 mV for the oxygen sensors. Click Connect to Oxygraph to establish communication with the instrument.
- After communication is established, a dialog box will open to name and save the experimental file. Save the file with the current date and calibration. After saving the file, a pop-up dialog box will appear for experimental details. No information is needed for the calibration run, and the box can be closed.
- Record oxygen concentration for at least 30 min to allow chambers to warm up and to record signals for air calibration. This can be done while preparing muscle fibers, as detailed in step 2 below.
- At the end of the calibration period, mark a calibration region across the region where the oxygen concentration (blue line) is stable. To do this, hold down the Shift key and the Left Mouse button and drag across a region on the timeline.
- Open the calibration window by going to the Oxygraph > O2 Calibration. For Air Calibration, select the mark generated in step 1.8. Click Calibrate and Copy to Clipboard.
- Repeat steps 1.6-1.8 for the remaining chamber. Perform air calibration daily. Stop the air calibration recording and save the file by disconnecting from the instrument. After the samples are ready, proceed to step 3 to conduct the assay.
2. Harvesting and permeabilization of skeletal muscle fibers
- Mouse tissue isolation
- For each sample to be analyzed, fill one well of a 12-well culture dish with 1 mL of BIOPS. Place plate on ice to chill.
- After euthanizing the mouse by carbon dioxide inhalation followed by cervical dislocation, harvest the skeletal muscle of interest, being sure to remove all connective tissue and fat (Figure 2A). Place the muscle in one of the wells containing BIOPS. Place on ice until fiber preparation.
NOTE: Any skeletal muscle can be examined using this protocol – the muscle type examined will be dependent on the experimental question one seeks to address. In the case of skeletal muscles with mixed fiber types, such as the gastrocnemius, it is possible to separate the tissue into white and red sections to measure predominantly fast twitch (white sections) and predominantly slow twitch (red sections) fibers (Figure 2B).
- Human tissue isolation
- Collect tissue following clinical protocol14. Quickly rinse the tissue with cold, sterile PBS and place it in a conical tube containing 2 mL of BIOPS solution. Keep tissue on ice until fiber preparation.
- Fiber preparation
- Permeabilization will take place in a 6-well tissue culture plate. Add 2 mL of BIOPS solution to one set of wells and 2 mL of MiR05 to another set of wells. One set of wells will be required for each sample being assayed.
- Pack a tray with ice and place a glass Petri dish upside down on the ice. Be sure to always keep the muscle on ice.
- Transfer harvested muscle samples to the chilled Petri dish platform using forceps. Using two fine-tipped forceps and a dissecting microscope with a light source, gently clean the tissue by removing any tendons, fascia, and adipose tissue that are attached to the muscle.
- After all extraneous non-muscle tissue has been removed, gently pull the muscle fibers apart with the fine-tipped forceps until they are small bundles of translucent fibers ranging from 0.75 – 1.0 mm in length (Figure 2C-D). After separating the fibers, use the forceps to transfer the fiber bundles to a well on the 6-well plate containing BIOPS solution on ice.
- Add 20 µL of 5 mg/mL saponin solution to each BIOPS well and incubate the plate on ice while gently rocking in a cold room for 20 min.
- After saponin treatment, transfer muscle fibers to the well containing MiR05 to rinse before the assay. Incubate on ice while gently rocking in a cold room for 15 min.
- After the fiber incubation in MiR05 is complete, gather the fibers with sharp forceps and gently blot the muscle fibers dry on a task wipe. Tare a 1.5 mL plastic microcentrifuge tube on a fine balance and place 2-3 mg of fibers in the tube. Record the final fiber weight on the side of each tube. Place the tube on ice. Repeat with other samples. Immediately proceed to step 3.
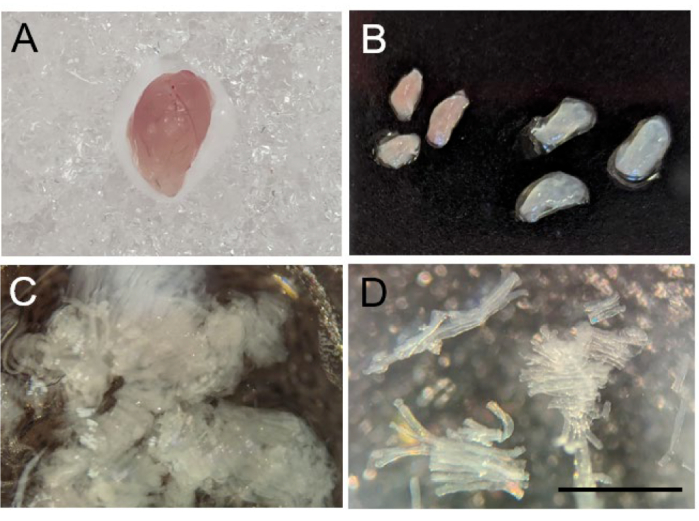
Figure 2: Separation of mouse skeletal muscle fibers. (A) Gross morphology of mouse gastrocnemius after harvest. (B) Dissection of gastrocnemius into red (left) and white (right) segments. (C) Mechanically separated muscle fibers. (D) A 10x image of successfully separated muscle fibers. Scale bar is 1 mm. Please click here to view a larger version of this figure.
3. Preparing muscle samples in the respirometer
- Vacuum out the MiR05 used for instrument calibration as detailed in step 1 above and add 2.1 mL of working MiR05 solution to each chamber.
- Initiate a new file for the experiment using the same instrument settings as detailed in step 1.5. above. Set the file name and save settings.
- Following setting the file name, the next screen dialog box will be for specific experimental details. Enter the sample information and the weight of each sample added to each chamber. Close the dialog box.
- Once in the new experimental file, set the calibration by opening the O2 Calibration window. Click Copy from file and select the file saved from step 1.9. above. Click the Calibrate and Copy to Clipboard button. Repeat the process for the remaining chamber.
- Using fine forceps, carefully transfer muscle fiber bundles into the respiration solution for the appropriate chamber. Repeat for the remaining chamber.
- Place the stoppers on the chamber and carefully semi-close the chamber by pushing the stoppers about halfway to the bottom. Once the o-rings on the stoppers engage with the chamber wall and there is resistance, use a twisting motion while pushing down to close. When the chamber is halfway closed, a small air bubble can be observed at the top of the chamber.
NOTE: Permeabilized fibers are subject to oxygen diffusion limitations under regular respiratory conditions. To circumvent this limitation, the assay is conducted under elevated oxygen concentrations11. - Fill a 10 mL plastic syringe with pure oxygen from an oxygen tank. Place the long, blunt needle supplied with the instrument on the syringe. Place the needle in the first chamber and slowly deliver approximately 1 mL of oxygen into the chamber.
- Monitor the chamber oxygen concentration in the respirometry software. When the chamber oxygen concentration reaches 350-400 nmol/mL, gently twist the stopper while pushing down to fully close the chamber. Look into the chamber and ensure that no air bubbles remain. If an air bubble is present, carefully and quickly reopen the chamber, add 100 µL of additional working MiR05 solution, and quickly close the chamber again. Repeat with the remaining chambers.
- Oxygen concentrations should be maintained above 250 nmol/mL during the assay. Add additional oxygen as needed by partially opening the chamber, adding more oxygen from the syringe into the air bubble, and carefully closing the chamber again.
- Normalize oxygen flux data to the mass of tissue used for the experiment. To adjust the graphs, change the layout to reflect O2 Flux (pmol O2 / (s x mg)). From the layout menu, select the 06 – Specific Flux per Unit Sample layout. Data will now be presented normalized to the amount of tissue in each chamber.
4. High resolution respirometry
- When the oxygen (blue line) concentration and O2 flux (red line) have stabilized following oxygen addition, begin the respiration protocol. Oxygen can be considered stable when the blue line is flat or slowly decreasing. O2 flux should also be flat and within 5 pmol O2 / s x mg.
- Add all reagents using glass syringes. It is important not to use the same syringe for substrates, inhibitors, and uncouplers. Have a separate syringe for each. After adding each compound, record oxygen flux for 1-2 min after the respiration rate stabilizes before adding the next reagent.
- Respiration measurement
- Using a 10 µL glass syringe, add 2.5 µL of 0.8 M malate to each chamber. Press F4 to mark the timeline and label the mark with M. Record stable O2 flux for 1-2 min.
- Add nutrient-specific reagents as described below.
- Aerobic Glycolytic: Using a 10 µL glass syringe, add 10 µL of 2 M glutamate and 5 µL of 2 M pyruvate to each chamber. Press F4 to mark the timeline and label the mark with G P. Record stable O2 flux for 1-2 min.
- Fatty acid: Using a 10 µL glass syringe, add 10 µL of 2 M glutamate and 10 µL of 10 mM palmitoyl-carnitine to each chamber. Press F4 to mark the timeline and label the mark with G PC. Record stable O2 flux for 1-2 min.
- Using a 25 µL glass syringe, add 20 µL of 0.5 M (with MgCl2) ADP to each chamber. Press F4 to mark the timeline and label the mark with ADP. Record stable O2 flux for 1-2 min.
- Using a 25 µL glass syringe, add 20 µL of 1 M succinate to each chamber. Press F4 to mark the timeline and label the mark with S. Record stable O2 flux for 1-2 min.
- Using a 10 µL glass syringe, add 5 µL of 4 mM Cytochrome C to each chamber. Press F4 to mark the timeline and label the mark with Cyt C. Record stable O2 flux for 1-2 min.
- Titrate in three 1 µL boluses of 1 mM FCCP using a 10 µL glass syringe. There is generally a mixing effect when adding FCCP that results in a brief decrease in O2 flux levels. Wait for the O2 flux to increase and stabilize before proceeding to the next step. Press F4 after each addition to mark the timeline and label mark with FCCP. Record stable O2 flux for 1 min after each addition.
- When the respiration assay is complete, gently remove stoppers by twisting and pulling upward. Rinse the chambers 3x with ultrapure H2O from a squirt bottle, followed by 3x with 70% ethanol from a squirt bottle. After the final ethanol rinse, fill the chambers with 70% ethanol for storage.
- Set the stoppers in the chambers until you feel resistance. Do not close the stoppers all the way. Place caps over the stoppers, save the assay file and disconnect the instrument from the Respirometry software. Turn off the instrument.
5. Data analysis
- Open the analysis file in the respirometry software. Extract data from stable oxygen flux regions obtained after injection of respiration compounds.
- To mark regions of interest, hold down the Shift key, click the Left mouse button, and drag the box across the stable O2 flux rate region for the assay stages detailed below.
- Assay stages14,15,16,17,18,19,20
- Mark the timeline after the addition of malate/glutamate/pyruvate (aerobic glycolytic protocol) or malate/glutamate/palmitoyl-carnitine (fatty acid protocol). This rate represents the Complex I State 2 respiration (LEAK(n)) rate.
- Mark the timeline after the addition of ADP. This rate represents Complex I State 3 respiration (CI OXPHOS) rate.
- Mark the timeline after addition of succinate. This rate represents Complex I+II State 3 respiration (CI+II OXPHOS) rate.
- Mark the timeline after adding Cytochrome C. This rate represents Complex I+II+Cytochrome State 3 respiration (CI+II+Cyt C OXPHOS) rate.
- Mark the timeline for the FCCP titration with the highest O2 flux rate. This rate represents Maximal Respiratory Rate (MAX ETS).
- Retrieve the data values for the marked regions on the timeline by selecting Mark > Statistics. Copy the O2 Flux rate (pmol O2/(s x mg)) for the marked chamber to a spreadsheet.
- Repeat marking and data extraction for the additional chambers.
- Calculate the respiratory control ratio (RCR) by dividing Complex I+II State 3 (following succinate addition) by Complex I State 2 (prior to ADP addition).
Figure 3 and Figure 4 show oxygen plots of aerobic glycolytic and fatty acid respirometry protocols, respectively, for properly prepared murine red and white gastrocnemius muscle fibers. Also shown are representative quantified results for reference. Figure 5 shows an oxygen plot of aerobic glycolytic respirometry in human muscle biopsy samples that were properly prepared. Representative quantified results are also shown. Note that for Figure 3, Figure 4, and Figure 5, addition of Cytochrome C after ADP addition does not produce an impact on oxygen flux, indicating that the outer mitochondrial membrane of the sample is intact. Figure 6 shows an oxygen plot of aerobic glycolytic respirometry where the addition of Cytochrome C after ADP results in a spike (40% increase) in oxygen flux, indicating the outer mitochondrial membrane has been damaged and thus the sample should not be used for respirometry – potential reasons for this result can be inappropriate handling or freezing/thawing of the tissue, prolong permeabilization of the tissue, and not using freshly isolated tissue.
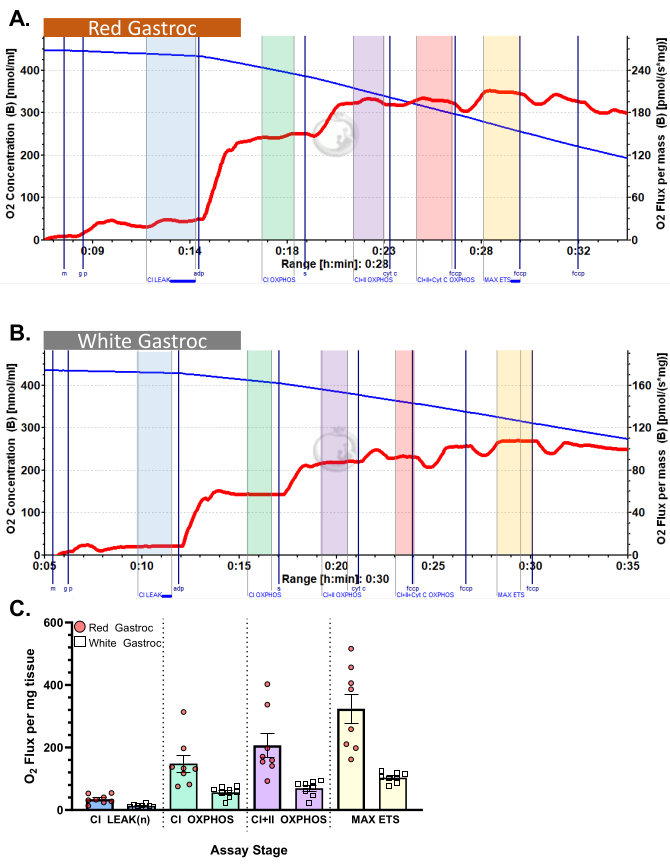
Figure 3: Oxygen consumption in mouse. The results show oxygen consumption in (A) red and (B) white gastrocnemius using pyruvate protocol. State 2 flux following the addition of malate, glutamate, and pyruvate (blue shade, CI LEAK). Significant stimulation of O2 consumption is observed after ADP administration (green shade, CI OXPHOS), with respiration driven further after the addition of succinate (purple shade, CI+II OXPHOS). Cytochrome C induced no significant increase (<15%), indicating the outer mitochondrial membrane is intact (orange shade, CI+II+Cyt C OXPHOS). Mitochondria are uncoupled following the addition of FCCP (yellow shade, MAX ETS). The blue line represents oxygen concentration in a closed chamber. The red line represents the rate of oxygen consumption (O2 flux). Compounds added: Malate (m), Glutamate (g), Pyruvate (p), Adenosine Diphosphate (ADP), Cytochrome C (cyt c), Carbonyl cyanide-p-trifluoromethoxyphenylhydrazone (FCCP). (C) The bar graph reflects representative results (n=8). Data are represented as ± SEM. Please click here to view a larger version of this figure.
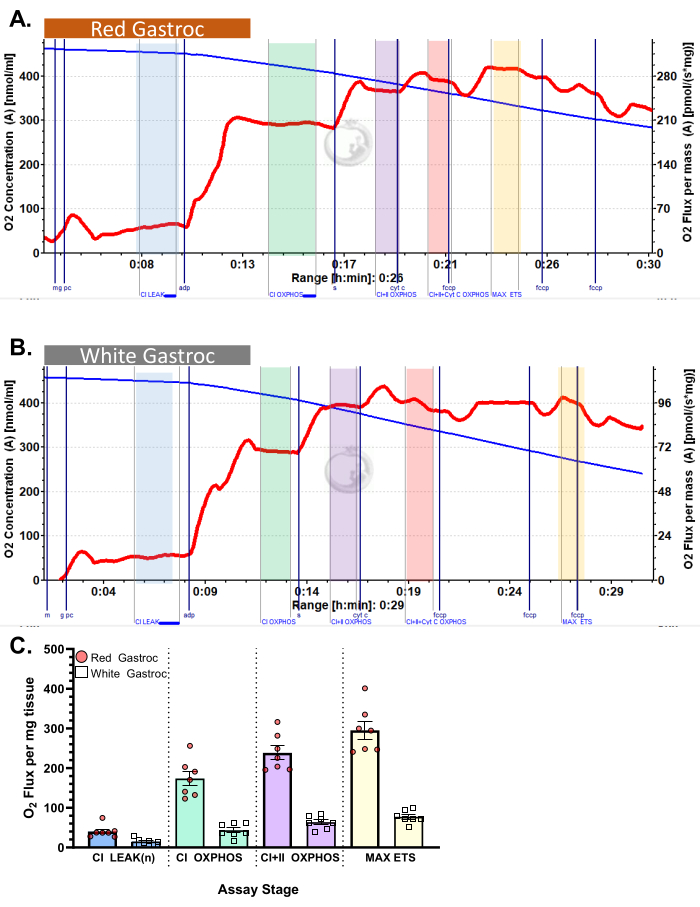
Figure 4: Oxygen consumption in mouse. The results show oxygen consumption in (A) red and (B) white gastrocnemius using palmitoyl-carnitine protocol. State 2 flux following the addition of malate, glutamate, and palmitoyl carnitine (blue shade, CI LEAK). Significant stimulation of O2 consumption is observed after ADP administration (green shade, CI OXPHOS), with respiration driven further after the addition of succinate (purple shade, CI+II OXPHOS). Cytochrome C induced no significant increase (<15%), indicating the outer mitochondrial membrane is intact (orange shade, CI+II+Cyt C OXPHOS). Mitochondria are uncoupled following the addition of FCCP (yellow shade, MAX ETS). The blue line represents oxygen concentration in a closed chamber. The red line represents the rate of oxygen consumption (O2 flux). Compounds added: Malate (m), Glutamate (g), Palmitoyl Carnitine (pc), Adenosine Diphosphate (ADP), Cytochrome C (cyt c), Carbonyl cyanide-p-trifluoromethoxyphenylhydrazone (FCCP). (C) The bar graph reflects representative results (n=7). Data are represented as ± SEM. Please click here to view a larger version of this figure.
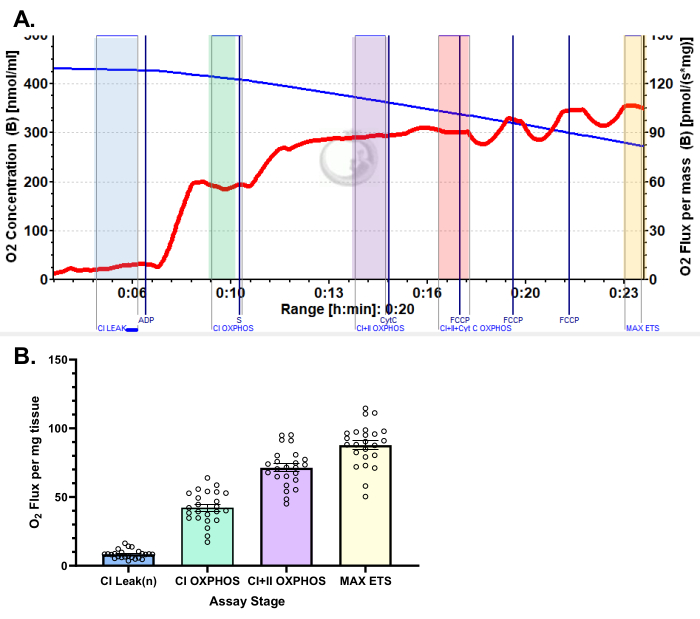
Figure 5: Representative results for oxygen consumption in human vastus lateralis using pyruvate protocol. (A) State 2 flux following the addition of malate, glutamate, and pyruvate (blue shade, CI LEAK). Significant stimulation of O2 consumption is observed after ADP administration (green shade, CI OXPHOS), with respiration driven further after the addition of succinate (purple shade, CI+II OXPHOS). Cytochrome C induced no significant increase (<15%), indicating the outer mitochondrial membrane is intact (orange shade, CI+II+Cyt C OXPHOS). Mitochondria are uncoupled following the addition of FCCP (yellow shade, MAX ETS). The blue line represents oxygen concentration in a closed chamber. The red line represents the rate of oxygen consumption (O2 flux). Compounds added: Malate (m), Glutamate (g), Pyruvate (p), Adenosine Diphosphate (ADP), Cytochrome C (cyt c), Carbonyl cyanide-p-trifluoromethoxyphenylhydrazone (FCCP). (B) The bar graph reflects representative results obtained from vastus lateralis biopsies (n = 24). Data are represented as ± SEM. Please click here to view a larger version of this figure.
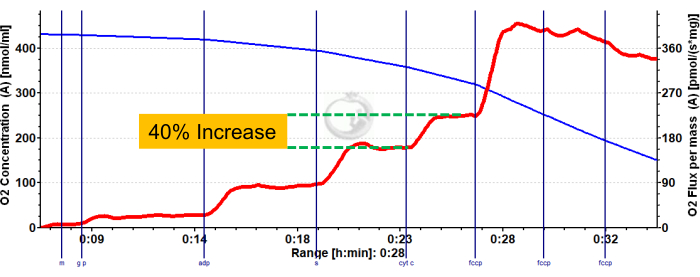
Figure 6: Representative result demonstrating compromised outer mitochondrial membrane integrity in mouse red gastrocnemius. State 2 flux following the addition of malate, glutamate, and pyruvate (blue shade, CI LEAK). Significant stimulation of O2 consumption is observed after ADP administration (green shade, CI OXPHOS), with respiration driven further after the addition of succinate (purple shade, CI+II OXPHOS). Cytochrome C induced a significant increase in O2 consumption (>15%), indicating damage to the outer mitochondrial membrane. The blue line represents oxygen concentration in a closed chamber. The red line represents the rate of oxygen consumption (O2 flux). Compounds added: Malate (m), Glutamate (g), Pyruvate (p), Adenosine Diphosphate (ADP), Cytochrome C (cyt c), Carbonyl cyanide-p-trifluoromethoxyphenylhydrazone (FCCP). Please click here to view a larger version of this figure.
Table 1: Reagent preparation of respiration compounds and respiration solutions. Details for preparing reagents necessary for the assay are presented, including final stock concentrations and how to prepare and store them. Please click here to download this Table.
