Ileal enteroids grown in ECM are analyzed visually and by qRT-PCR for their relative health status and differentiation states as a means of quality control for ileal enteroid cultures and for use in monolayers. Undifferentiated ileal enteroids grown in ECM appear clear and cystic in morphology, indicating the presence of many stem cells (Figure 1A). Over time, undifferentiated ileal enteroids grown in growth media may take on an intermediate phenotype where some will appear cystic and some appear opaque (Figure 1B). Frequently, our undifferentiated samples resemble those shown in Figure 1B rather than Figure 1A. These intermediate cultures contain more terminally differentiated enterocytes as measured by expression of the enterocyte marker, sucrase isomaltase (SI), and presumably extruded dead enterocytes in the lumen contribute to their dense appearance. Ileal enteroids can be used in this intermediate state for monolayer development, but it must be kept in mind that the quantity of intestinal stem cells present in the cultures may be low, and some differentiated cell types may be present (for example, see qRT-PCR levels in undifferentiated samples grown in ECM resembling Figure 1B di Figure 2). For comparison, ileal enteroids cultured with differentiation media in ECM for 5+ days will appear uniformly darkened and lobular and cultures with this morphology are not good candidates for seeding monolayers (Figure 1C).
Expression of stem cell genes and genes of intestinal cell differentiation can be analyzed by qRT-PCR as another means to assess the health status of ileal enteroids grown in ECM and their differentiation capabilities once seeded as monolayers on transwells. The expression of a stem cell gene, LGR5, an enterocyte gene, SI, a goblet cell gene, MUC2, and a Paneth cell gene, LYZ, is compared between undifferentiated ileal enteroid cultures grown in ECM and differentiated ileal enteroid monolayers in the presence or absence of RANKL/TNFα (Figure 2). While the values may differ between experiments, expression of LGR5 should decrease after differentiation of monolayers18,26. LGR5 expression is usually not detected in the differentiated ileal monolayers without RANKL and TNFα by day 7. Conversely, expression of markers of differentiation of specific cell types, such SI and MUC2, increase after differentiation18. Expression of LYZ generally decreases after differentiation in our cultures. If the ileal enteroid cultures used for making monolayers look more like Figure 1B than Figure 1A, increases in intestinal differentiation markers may be modest after differentiation because these initial cultures are heterogeneous in intestinal cell types and have a higher basal level of SI and MUC2. However, differentiation in monolayers still occurs as assessed by loss of LGR5 expression and microscopy (see below). Furthermore, addition of RANKL and TNFα to the differentiation media reduces the loss of LGR5 expression (Figure 2). In parallel, the expression of SI and MUC2 are slightly lower than in the differentiated condition lacking RANKL and TNFα although their levels increase above the undifferentiated condition.
M cell differentiation in monolayers is determined both by qRT-PCR and immunofluorescence using two M cell specific markers including cell surface glycoprotein 2 (GP2) and transcription factor SpiB21. Expression of GP2 and SPIB is upregulated in the ileal enteroid-derived monolayers in the presence of RANKL and TNFα and is not detected in non-RANKL and TNFα treated samples (Figure 3). Expression of these markers can also be normalized to a piece of small bowel tissue22, if available. This permits the fold change of these M cell markers to be compared to tissue that has M cells rather than to control monolayers that have no expression of these markers and allows standardization between experiments in one lab. M cells are also detected by surface expression of GP2 by immunofluorescence (Figure 4). Typically, in a confluent monolayer, 1 to 5 M cells are observed in a given microscope field at 40X magnification by days 6 through 8 post-seeding in samples treated with RANKL and TNFα (Figure 4A-D). No GP2 expression is seen in the untreated samples (Figure 4E). The orthogonal view of the XZ plane overlaid with a phalloidin probe shows actin structures surrounding each cell and GP2 expression on the apical surface of M cells (Figure 4F-G). This model recapitulates the low frequency of M cells found in the human intestine1,2,8. To purify and isolate M cells for further study, M cells can be stained using GP2 surface expression and sorted using FACS for GP2+ cells.
M cells bind to and transport antigen from the intestinal lumen to the immune cells residing beneath the epithelium2. Secretory IgA produced in the intestine binds to bacteria and can bind to the apical surface of M cells to facilitate transport of the microbes27,28. To determine if the M cells developed in this model are able to bind to IgA, human serum IgA is added to the upper chamber, allowed to bind for 1 h, and then the monolayers are prepared for immunofluorescence analysis. The presence of IgA on M cells is visualized using a fluor-conjugated secondary antibody that recognizes the heavy chain of human serum IgA. M cells treated with IgA for 1 h have IgA bound to the apical surface (Figure 5A), whereas M cells in control wells that were only treated with the secondary antibody to IgA have no detectable signal (Figure 5B). Further, IgA specifically binds to the apical surface of M cells and is not found bound to any cells lacking GP2 surface stain. In addition, M cells have characteristically shorter dense actin on their apical surface2. To analyze M cell morphology in this model, ileal enteroid-derived monolayers are grown for 7 days and harvested for immunofluorescence analysis of F-actin using phalloidin. Measurements of actin pixel intensity are calculated for M cells and for non-M cells that are directly adjacent to each M cell using ImageJ software (Figure 6A). Actin intensity is reduced on GP2+ M cells in this model and a representative image is shown in Figure 6B. Overall, the M cells developed in this ileal enteroid-derived monolayer model have characteristic gene expression, morphology and some M cell functions of human intestinal M cells, such as binding to IgA.
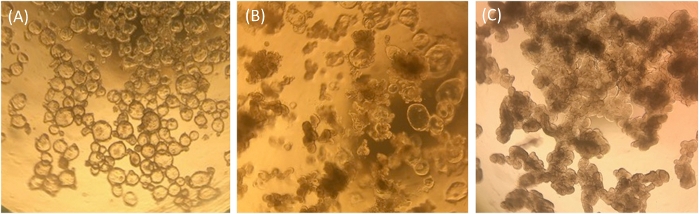
Figure 1: Representative morphology of human ileal enteroids in ECM one-week post-splitting. (A) Clear and cystic undifferentiated ileal enteroids. (B) Intermediate phenotype with some cystic ileal enteroids and some opaque lobular ileal enteroids. (C) Darkened and lobular differentiated ileal enteroids. Images taken through the lens of an optical light microscope at 4x magnification using an iPhone7 camera. Please click here to view a larger version of this figure.
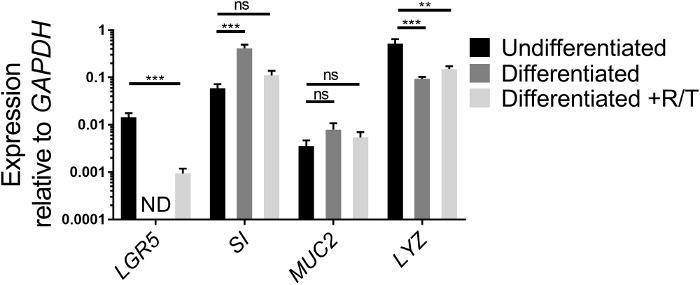
Figure 2: Relative expression of stem cell and differentiation markers of human ileal enteroids grown in ECM or differentiated as monolayers. Ileal enteroids were grown for 7 days in ECM (Undifferentiated) or grown and differentiated as monolayers without (Differentiated) or with RANKL and TNFα (Differentiated +R/T). Ileal enteroid cultures or monolayers were harvested in Trizol for RNA extraction. Gene expression was determined by qRT-PCR and is expressed relative to GAPDH. Data is average of 3 independent wells of ileal enteroids or monolayers per condition. Error bars indicate SEM. ND is not detected. Statistical significance was determined on log-transformed values using one-way ANOVA with Dunnett's multiple comparisons test comparing to the Undifferentiated. ** p < 0.01, *** p < 0.001 Please click here to view a larger version of this figure.
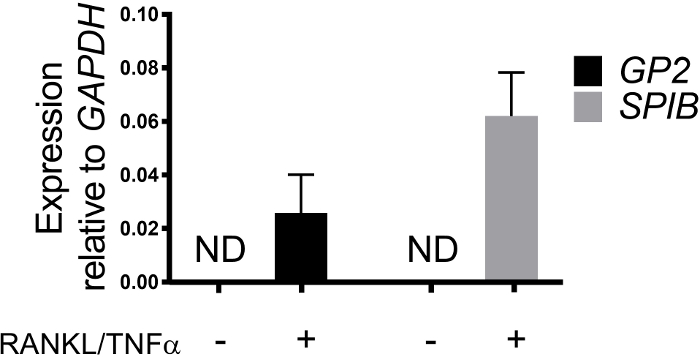
Figure 3: Relative expression of M cell specific markers GP2 and SPIB from human ileal enteroid-derived monolayers. RANKL/TNFα treated and non-treated human ileal enteroid-derived monolayers were harvested in Trizol for RNA extraction after 7 days post-seeding. Gene expression was determined by qRT-PCR and is expressed relative to GAPDH. Data is average of 6 independent monolayers per condition. Error bars indicate SEM. ND is not detected. Please click here to view a larger version of this figure.
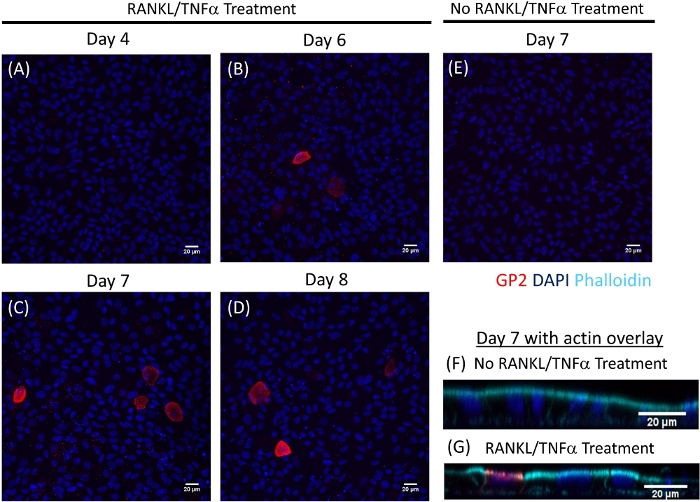
Figure 4: Immunofluorescence of surface GP2 expression on M cells in human ileal enteroid-derived monolayers over time. RANKL/TNFα treated and non-treated human ileal enteroid-derived monolayers were fixed in 4% PFA and stained for immunofluorescence on various indicated days post-seeding. Images were analyzed using ImageJ software. DAPI = Blue; Glycoprotein 2 (GP2) = Red. (A-D) RANKL/TNFα treated monolayers at various days post-seeding. (E) Non-treated monolayer harvested at day 7 post-seeding. (F–G) Orthogonal XZ plane of monolayers at day 7 post-seeding overlaid with phalloidin probe for F-actin. Phalloidin = Cyan. Please click here to view a larger version of this figure.
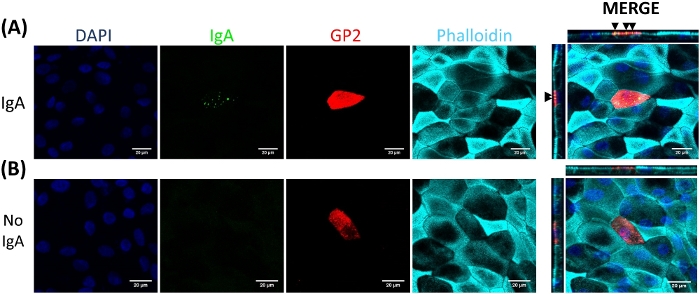
Figure 5: IgA binds specifically to the apical surface of M cells. RANKL/TNFα-treated human ileal enteroid-derived monolayers were grown for 7 days and then (A) treated with 10 μg of human serum IgA for 1 h or (B) mock-treated with PBS only (No IgA control). After 1 h, monolayers were washed 2x in PBS, were fixed in 4% PFA, permeabilized with 0.1% TritonX-100, and stained for immunofluorescence. Images were analyzed using ImageJ software and are representative of 3 independent experiments. DAPI = Blue; Glycoprotein 2 (GP2) = Red; Antibody to human serum IgA = Green; Phalloidin = Cyan. Black arrows denote IgA bound to apical surface of M cell. Please click here to view a larger version of this figure.
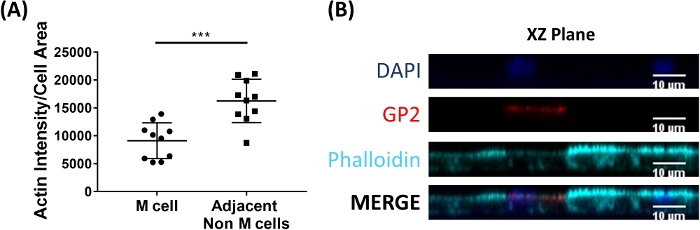
Figure 6: M cells have reduced actin intensity compared to adjacent non-M cells. RANKL/TNFα-treated human ileal enteroid-derived monolayers were grown for 7 days and then fixed in 4% PFA and were stained for immunofluorescence. (A) Using ImageJ, GP2+ M cells were outlined using the Freehand Selection Tool and measurements of Area and Integrated Density were taken in the Phalloidin channel. The same analysis was then completed for each adjacent non-M cell that neighbors the M cell. The Raw Integrated Density was divided by the Area of each individual cell for normalization. The average Integrated Density/Area was calculated for each adjacent non-M cells for each M cell. Images were analyzed from 3 independent experiments; each dot is an M cell or average of neighboring cells. Error bars indicate SD. Statistical significance was determined on log-transformed values using a Paired t test. *** p = 0.0001 (B) Representative image of XZ Plane from plot in A. Images were analyzed using ImageJ software. DAPI = Blue; Glycoprotein 2 (GP2) = Red; Phalloidin = Cyan. Please click here to view a larger version of this figure.
