For these experiments, four eleven-week-old female wild type C57/Black6 mice were used. One mouse was used to conduct a full cerebellar dissection which is referred to as 'bulk cerebellum' and allowed for the comparison of RNA levels in dissected regions to a full dissection. The other three mice were used to conduct the cerebellar dissection described in this protocol. Using three mice makes it possible to ensure that the trends detected in the levels of RNA are reproducible across mice.
Figure 1A represents the mouse cerebellum – vermis in blue and hemispheres in yellow. Sagittal schematics of the vermis (anterior regions in dark blue, posterior regions in light blue) and hemisphere (in yellow). The DCN are highlighted in red dotted boxes. A successful dissection will start with the initial razor blade placement down the midline of the brain (Figure 1B); this guides the successful placement of the following six blades (Figure 1C). This leaves six sagittal sections (Figure 1D), four lateral/hemisphere sections (outlined in purple) and two midline/vermal sections (outlined in light blue). The 4 lateral sections contain the DCN; Figure 1E depicts a successful DCN punch dissection. The midline vermal sections will most likely have half of the most medial DCN present in the 1 mm thick section, however it will not be present all the way through and isn't possible to reproducibly dissect out. The midline vermal sections are separated into anterior and posterior lobules depicted in Figure 1F. A successful dissection sets up the rest of the experiments for success.
Real Time qualitative Polymerase Chain Reaction (RTqPCR) results demonstrate the feasibility of assessing the gene expression levels of each individual region as well as serve to validate dissections. We used primers detecting genes that show gradient expression from anterior to posterior cerebellar cortex and enrichment in the DCN and compared their expression to the bulk cerebellar lysates.
We assessed the expression levels of three genes: aldolase C, parvalbumin, and Kcng4. Aldolase C, also known as zebrin II, is an enzyme that is expressed in a consistent banding pattern through the cerebellum. It is expressed more highly in the posterior vermis than the anterior vermis. There are also bands in the hemispheres13,14. Parvalbumin which is a calcium binding protein that is expressed in inhibitory cells. Based on the Allen Brain Atlas, parvalbumin seems relatively uniformly expressed throughout the cerebellar cortex and in the DCN (http://mouse.brain-map.org/gene/show/19056). Kcng4, a potassium voltage gated channel subunit, appears to be enriched in the DCN and in the anterior compared to the posterior lobes (https://mouse.brain-map.org/gene/show/42576). Quantitative expression analysis showed that, as expected, aldolase C is more highly expressed in the posterior cerebellar vermis (CCpV) but lower in the DCN and the anterior region of the vermis (CCaV) when compared to the bulk cerebellar dissection (Figure 2A). Parvalbumin is similarly present in the DCN, anterior vermis, posterior vermis, and hemisphere cerebellar cortices as in the bulk cerebellar extracts (Figure 2B). Kcng4 is significantly enriched in the DCN and the anterior vermis (CCaV) and it is not significantly enriched in the posterior vermis (CCpV) or hemispheres (CCH) when compared to the bulk extraction (Figure 2C). This result follows what was expected based on the pattern seen in the Allen Brain Atlas. Thus, gene expression analysis validates the dissection protocol and confirms that good quality RNA can be obtained and tested.
To directly compare the expression of aldolase C across the cerebellar cortex, the expression levels were compared to where it is supposed to be lowest, the anterior vermis (CCaV) (Figure 3). The expression level of aldolase C was significantly higher in the posterior vermis (CCpV), and trending higher in the cerebellar hemispheres (CCH), though not quite significantly so. This trend in the cerebellar hemispheres is likely because there are bands of aldolase C in the hemispheres, and the dissection captures aldolase negative and positive bands.
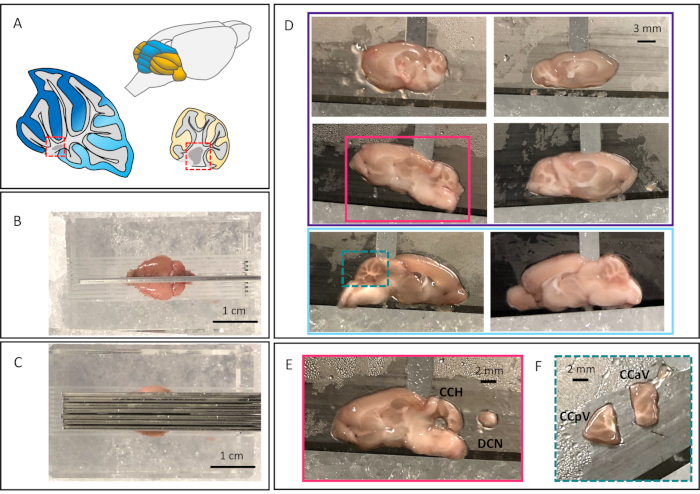
Figure 1: Representative images of cerebellar dissection
A. Schematic of mouse cerebellum with vermis in blue and hemispheres in yellow. Sagittal cerebellar schematics of both vermis and hemisphere. Vermis is in blue, with dark blue marking the anterior vermis, and light blue marking posterior vermis. Hemisphere in yellow. The DCN are marked in each with red-dotted boxes. B. Full brain in sagittal mouse brain matrix, with razor blade down midline. C. Placement of three razor blades 1 mm apart on either side of the midline. D. The resulting six sagittal brain sections. Four contain lateral/hemisphere cerebellar sections (the top four images outlined in purple). Two contain medial/vermal cerebellar sections (bottom two images, outlined in light blue). E. Representative lateral/hemisphere cerebellar section with DCN punch dissected to the right of the section (section outlined in pink in Figure 1D). F. Cerebellar section in Figure 1D with square dotted turquois around it, dissected into anterior and posterior vermal lobules. Please click here to view a larger version of this figure.

Figure 2: Relative gene expression in isolated specific regions of the cerebellum.
Relative expression of aldolase C (2A), parvalbumin (2B), and Kcng4 (2C) normalized to Rps 18 (protein associated with ribosomal RNA expressed in all cells), and using bulk cerebellar extract as a reference. As expected, aldolase C expression was higher in the posterior vermis and lower in the DCN and anterior vermis (2A). As expected, based on Allen Brain Atlas, parvalbumin expression is uniformly expressed across each extracted region, while Kcng4 expression is significantly enriched in the DCN and CCaV. One-way ANOVA, with a Tukey's post hoc test. *p<.0.005, ** p<.0.0001 relative to the bulk cerebellar extract. Histograms represent average values for N=3 with values for each individual mouse shown as dots. Error bars represent standard error of the mean. DCN (Deep cerebellar nuclei), CCaV (the cerebellar cortex of anterior vermis), CCpV (the cerebellar cortex of the posterior vermis), and CCH (the cerebellar cortex of the hemispheres). N=3 mice for cerebellar dissection regions. N=1 bulk extract. Experiment done in triplicate. Please click here to view a larger version of this figure.
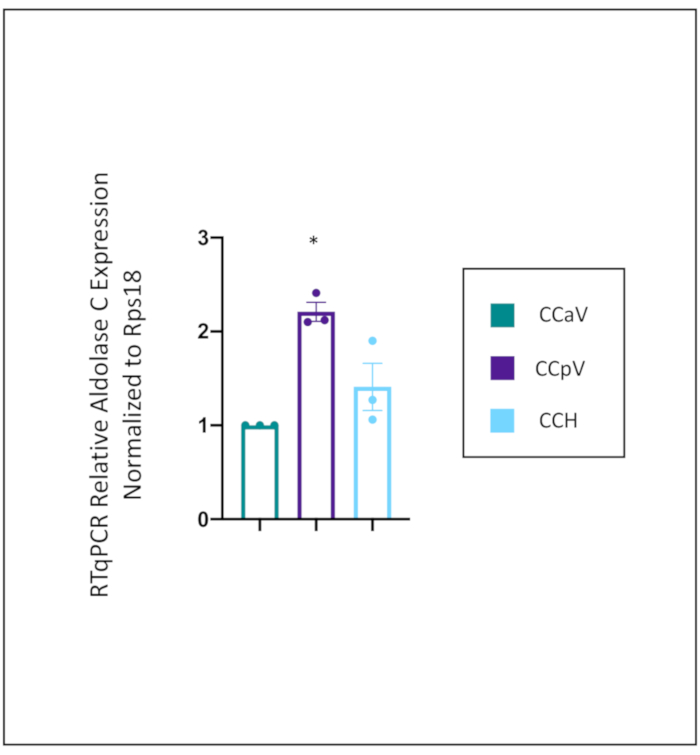
Figure 3: RTqPCR Relative gene expression of aldolase C across cerebellar cortex
Relative expression of aldolase C in specific regions of the cerebellar cortex – anterior vermis (CCaV), posterior vermis (CCpV), and hemispheres (CCH). Gene expression level of aldolase C was normalized to Rps18 and compared to the expression level in the anterior vermis. As expected aldolase C expression was enriched in the posterior vermis. One-way ANOVA, with a Tukey's post hoc test. *p<.0.005 relative to CCaV. Error bars represent standard error of the mean. CCaV (the cerebellar cortex of anterior vermis), CCpV (the cerebellar cortex of the posterior vermis), and CCH (the cerebellar cortex of the hemispheres). N=3 mice, experiment done in triplicate. Please click here to view a larger version of this figure.
