1. Bacterial isolate collection
- Obtain P. aeruginosa isolates from a cohort of pediatric patients with CF undergoing eradication treatment with inhaled tobramycin at SickKids (Toronto). Freeze isolates at -80 °C in glycerol citrate and sub-culture at least three times prior to use.
2. In vitro biofilm formation
NOTE: Use a chambered coverglass method1 for in vitro biofilm formation with modifications. The overall workflow of this model is shown in Figure 1.
- Grow P. aeruginosa isolate overnight at 37 °C on blood agar prepared with tryptic soy agar and 5% sheep blood (see Table of Materials).
- Inoculate 1–2 bacterial colonies from the blood agar into 4 mL of lysogeny broth (LB). Grow overnight at 37 °C on a shaker set to 225 rpm.
- Prepare a 1:100 dilution of overnight inoculum by adding 40 μL of the culture in 4 mL of fresh LB. Grow for 3–4 h at 37 °C on a shaker set to 225 rpm, to achieve an optical density of approximately 0.1 at 600 nm (OD600) (early log phase).
- Transfer 220 μL of the inoculum to each well of an 8-chambered coverglass slide. Incubate undisturbed at 37 °C for 24 h.
- Slowly remove the medium from each well to prevent the biofilms from detaching at the base.
NOTE: Tilt the slide forward at a 45° angle and aspirate the medium from the bottom corners of each chambered well without touching the base with pipette tip. - Prepare and slowly add 100 μL of 56 μg/mL fluorescent labelled monoclonal antibody (mAb) (see Table of Materials) to the side of the designated chambered wells. Incubate at room temperature (RT) for 1 h to allow antibody attachment to the bacterial antigen epitope.
NOTE: Prior to use, dilute the fluorescent labelled (red) mAb, Psl0096, in LB to obtain a final concentration of 56 μg/mL. Psl0096 is an anti-Psl mAb (optimized affinity derivative of Cam003), which binds to the class I Psl epitope—a key EPS matrix component of P. aeruginosa biofilms involved in initial cell attachment and structural integrity15. - Prepare and slowly add 100 μL of a 1000 μg/mL antibiotic solution (see Table of Materials) to the side of the designated chambered wells. Incubate undisturbed at 37 °C for 24 h.
NOTE: Prior to use, dilute a 50 mg/mL stock of tobramycin antibiotic in LB to obtain a final concentration of 1000 μg/mL.
3. Biofilm fluorescent staining
- Prepare a 0.01 mM solution of a live-cell-staining fluorescent dye. Slowly remove the medium from the chambered wells and add 200 μL of the dye mixture to each well. Incubate at RT in the dark for 45 min.
NOTE: Prior to use, prepare the live cell staining (green) fluorescent dye by adding 4 μL of a 5 mM stock to 2 mL of LB. - Slowly remove the medium from each well, and wash 2x with 200 μL of fresh LB.
- Add 200 μL of fresh LB to each well and proceed to examination via confocal microscopy.
4. Image acquisition by confocal microscopy
NOTE: The image-processing and COMSTAT analysis procedure is presented in Figure 2. Acquire images of wells on the same day of biofilm staining. If delay in visualization exceeds 1 h, refrigerate the chambered coverglass in the dark until further processing.
- Acquire images of wells using a confocal microscope system (see Table of Materials) with appropriate laser excitation wavelengths and filter sets for acquisition.
NOTE: Here, excite the fluorescent labeled (red) mAb and live cell (green) stain using 561 and 491 nm excitation wavelengths, respectively. - Capture layered z-stack images (from the substratum to the top of each biofilm segment) in increments of 0.3 μm with a 20–25x water immersion lens. Take a at least 6 image stacks per well.
NOTE: Here, visualize the images using a high-resolution camera with a 25x water immersion lens and processed using image analysis software (see Table of Materials). Keep software setup and digital imaging parameters (i.e., brightness and sensitivity) constant for all acquisitions in a single experiment. - Save images as OME-TIFFs for COMSTAT analysis.
NOTE: Ensure that OME-TIFFs are saved separately for each channel (i.e., red and green). This step varies depending on the image analysis software used. - Repeat steps 2.1–4.3 to capture images from a total of 3 biological replicates (i.e., 3 independent experiments) per bacterial isolate.
5. COMSTAT analysis
NOTE: Analyze images quantitatively using the freely available computer program, COMSTAT16,17, rewritten as a plugin (Comstat2) to ImageJ. Read the general instructions for analyzing image stacks of biofilms within the downloaded package. This contribution provides a summarized protocol, with selected ImageJ processing steps and COMSTAT features recommended for quantifying the effects of antimicrobials on biofilm formation.
- Download the Comstat2 package from http://www.comstat.dk/. Within the installed folder, locate ImageJ and run it.
- Create a source folder on desktop and add a single OME-TIFF to the folder.
- Open OME-TIFF from the source folder, and delete any empty layers containing no biomass. These layers will be either the first or last few layers of the z-stack.
NOTE: Microscope-defined z-stack boundaries are sometimes overestimated by users. Deleting these empty layers establishes a more refined z-stack boundary for COMSTAT analysis. - Import OME-TIFF in ImageJ by selecting File | Import | Image Sequence. Locate the source folder, highlight without opening it, and click Select. A ‘Sequence Options’ window will appear. Select OK.
NOTE: To import additional images in ImageJ, first remove the previous OME-TIFF from the source folder, then add the new OME-TIFF to the folder, and repeat steps 5.3 and 5.4. - Flip the orientation of the biofilm by selecting Image | Transform | Flip Z to position the substratum as the first (topmost) stack.
NOTE: COMSTAT algorithms read biofilms in the z-direction from top (stack 1) to bottom. Depending on the confocal microscope system used, the OME-TIFF output can be inverted. Thus, it is important to reverse the order of slices by positioning the substratum as image stack 1 to prevent output data from becoming flawed. - Define image properties by selecting Image | Properties. A ‘Source’ window will appear.
- Specify ‘Unit of Length’ as ‘micron’.
- Mathematically determine ‘Pixel width’ and ‘Pixel height’ using the following equation:
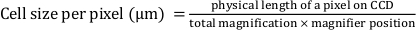 (1)
(1)
Here, ‘Pixel width’ and ‘Pixel height’ are defined as 0.427, where the physical length of a pixel on the charge-coupled device (CCD) is 16 μm; total magnification is 25x; and magnifier position is 1.5x.
NOTE: The equation used to calculate cell size per pixel may vary depending on the microscope camera manufacturer. Alternately, ‘Pixel width’ and ‘Pixel height’ can be defined by spatial calibration (refer to https://imagej.net/Spatial_Calibration). - Define ‘Voxel depth’ as 0.3 (i.e., incremental space between each z-stack layer). Select OK in the ‘Source’ window.
- Adjust the image threshold by selecting Image | Adjust | Threshold. A ‘Threshold’ window will appear.
NOTE: Objects in the image will appear red with a greyscale background. Alternatively, threshold can be adjusted in black-and-white, by using the drop-down menu in the ‘Threshold’ window to change ‘Red’ to ‘B&W’.- In the image window, adjust the slider to the far right (i.e., topmost layer of the biofilm). To remove background noise, use the ‘Threshold’ window, which displays a histogram of the image, to manually set the maximum and minimum threshold values. First, set the maximum threshold value by adjusting the lower slider as far right as possible. Second, use the upper slider to adjust the minimum threshold value, which segments the image into two separate phases: red biomass and greyscale background (Figure 3).
NOTE: The in vitro biofilm formation and fluorescent microscopy procedure described herein generates OME-TIFFs in the ideal case, allowing for images to be segmented into two distinct phases by means of a simple histogram threshold method. However, in some cases, the histogram distinction between the different phases is not as clear. This can be due to the presence of extensive background noise, varying background intensities, or a low intensity contrast between biomass and background. In such cases, users should adopt an enhanced segmentation procedure18,19.- Alternatively, adjust thresholds algorithmically for individual images using the left drop-down menu set as ‘Default’ in the ‘Threshold’ window. This feature provides 17 different algorithmic threshold options to choose from (refer to https://imagej.net/Auto_Threshold). Select the most applicable option, then ‘Auto’ to set threshold.
- When threshold values are adjusted, use the slider in the image window to scroll through each layer to ensure that background noise is sufficiently removed throughout.
- Select Set in the ‘Threshold’ window to first fix the lower threshold value. A ‘Set Threshold Levels’ window will appear. Select OK. Select Set again and repeat this step to fix the maximum threshold value.
NOTE: Each time ‘Set’ is selected, the lower slider may adjust automatically. In such cases, manually re-adjust the slider (or select ‘Auto’ if using one of the algorithmic thresholds) and repeat steps 5.6.2–5.6.3. The main idea is that whenever the sliders are re-adjusted automatically, ‘Set’ should be selected an additional two times afterwards to ensure that both the upper and lower thresholds are fixed. - Select Apply and a ‘Convert Stack to Binary’ window will appear. Select OK, and then exit the ‘Threshold’ window.
- Save the newly adjusted OME-TIFF by selecting Plugins | Bio-Formats | Bio-Formats Exporter|. Enter a new file name and save as OME-TIFF in the source folder. A ‘Bio-Formats Exporter – Multiple Files’ window will appear. Select OK. A ‘Bio-Formats Exporter Options’ window will appear. Select OK.
NOTE: Ensure that only the newly adjusted, black-and-white OME-TIFF(s) are saved in the source folder before proceeding to COMSTAT analysis. Remove all original OME-TIFFs from the folder.
- In the image window, adjust the slider to the far right (i.e., topmost layer of the biofilm). To remove background noise, use the ‘Threshold’ window, which displays a histogram of the image, to manually set the maximum and minimum threshold values. First, set the maximum threshold value by adjusting the lower slider as far right as possible. Second, use the upper slider to adjust the minimum threshold value, which segments the image into two separate phases: red biomass and greyscale background (Figure 3).
- Run COMSTAT by selecting Plugins | Comstat2 |. An ‘About’ window will appear. Select OK. Three windows will appear.
- In the ‘Observed Directories’ window (top right), select Add. Locate the source folder, highlight without opening it, and select Choose. An ‘Images in Directories’ window will appear (top left) that lists the OME-TIFFs to be analyzed via COMSTAT.
- On the ‘Comstat 2.1’ window (bottom right), de-select ‘Automatic thresholding (Otsu’s method)’ to ensure that the software uses threshold values previously set up for individual OME-TIFFs. Also, de-select Connected Volume Filtering (CVF) to ensure that very thin parts of the biofilm as well as free floating cells or biomass found within voids of the biofilm structure are included in the analysis.
NOTE: Here, de-select CVF because COMSTAT analysis is performed on very early biofilms (24 h initial growth) and remaining planktonic cells/colonies post antimicrobial treatment. For mature biofilms, select CVF to ensure that only biomass connected to the biofilm structure is quantified. - On the ‘Comstat 2.1’ window (bottom right), select desired features for quantitative analysis. Here, select Bio Mass, Thickness Distribution, and Surface Area. Select Go to run the program. In the ‘Log’ window (bottom left), the output data are shown processing until ‘Done with selected functions/images!’ appears. Record the COMSTAT measurements. These measurements are also automatically saved as TXT files in the source folder.
A P. aeruginosa isolate cultured from an infected patient with CF is used to demonstrate the strengths of this approach in accurately quantifying antimicrobial-induced changes in in vitro biofilm architecture. The overall workflow of this model is represented in Figure 1. The image-processing and COMSTAT analysis procedure in ImageJ is shown in Figure 2. A simple histogram thresholding approach for image segmentation in ImageJ, applied to a CLSM z-stack image (saved as OME-TIFF), is shown in Figure 3. Biofilm structural changes caused by tobramycin and anti-Psl mAb, Psl0096, is shown in Figure 4. Representative confocal images of biofilm segments separated into live cell (green) and antibody (red) channels are shown in Figure 4A. Corresponding data of three COMSTAT parameters, including average biofilm thickness, biomass, and surface-to-biovolume ratio are shown in Figure 4B-D. Overall, COMSTAT data demonstrate significant differences among biofilm structures compared to control wells. With tobramycin, a clear reduction in average thickness and biomass can be observed in Figure 4B and Figure 4C, respectively. However, in the presence of the anti-Psl mAb, Psl0096, the P. aeruginosa biofilm is resistant to tobramycin. When examining surface-to-biovolume ratio (Figure 4D), a significant reduction is observed with anti-Psl mAb, Psl0096 (p < 0.0001), in both tobramycin-exposed and unexposed biofilms, representing the formation of aggregates (Figure 4A).
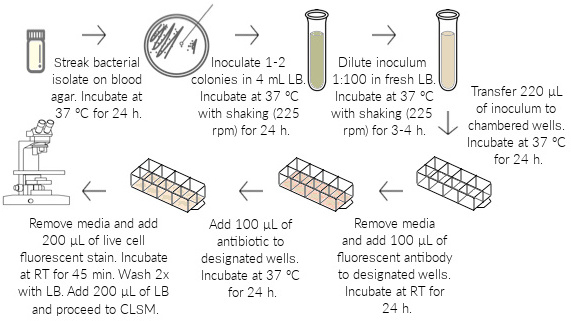
Figure 1: In vitro biofilm formation. A Pseudomonas aeruginosa isolate is grown on blood agar and inoculated in LB overnight. The overnight culture is diluted 1:100 and grown to early log phase (3–4 h incubation) to obtain a final OD600 of approximately 0.1. The OD-adjusted inoculum is seeded into each well of an 8-chambered coverglass and grown for 24 h, allowing biofilms to attach and mature at the base of the wells. Media is carefully removed from the wells, and a fluorescent labelled (red) mAb is added to the designated wells. The chambered coverglass is incubated for 1 h, allowing the antibody to bind to the bacterial antigen epitope. The antibiotic is added to designated wells, and biofilms are grown for 24 h. Medium is removed from the wells, and a live-cell fluorescent (green) stain is added. Following 45 min of incubation, the wells are washed 2x with fresh LB, and confocal imaging is performed in LB.
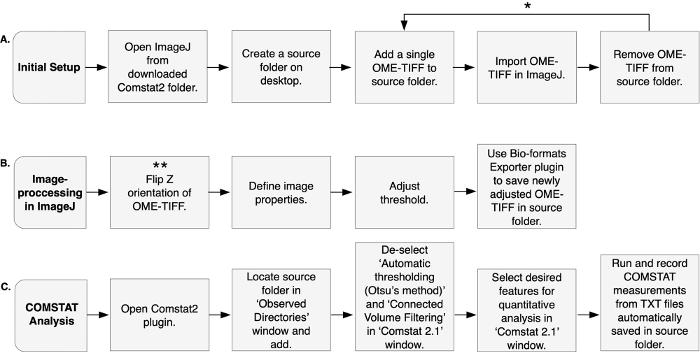
Figure 2: Image-processing and COMSTAT analysis. Comstat2, as a plugin to ImageJ, is used for the quantitative analysis of in vitro biofilm architecture. (A) Initial setup is performed by first opening the ImageJ program found within the downloaded Comstat2 package. It is then recommended to create a source folder, from which a single OME-TIFF can be added and imported into ImageJ. Once imported, the OME-TIFF is deleted from the source folder. * These steps can be repeated to add additional OME-TIFFs, allowing the operator to process multiple images at a time. (B) Image-processing is performed entirely in ImageJ. The orientation of OME-TIFF is flipped in the z-direction, assigning the first layer of biofilm substratum as stack 1. ** This step is only necessary if the OME-TIFF output from the confocal microscope is inverted. Image properties are defined, and the threshold of OME-TIFF is adjusted manually. The Bio-formats Exporter plugin is used to save the newly adjusted OME-TIFF in source folder. (C) COMSTAT analysis is performed using the Comstat2 plugin. The source folder containing the OME-TIFF(s) is added. Specific features that may flaw results are de-selected, while desired features for quantitative analysis are selected. ‘Run’ is selected to initiate analysis. Once completed, measurements are recoded from TXT files automatically saved in the source folder. Please click here to view a larger version of this figure.
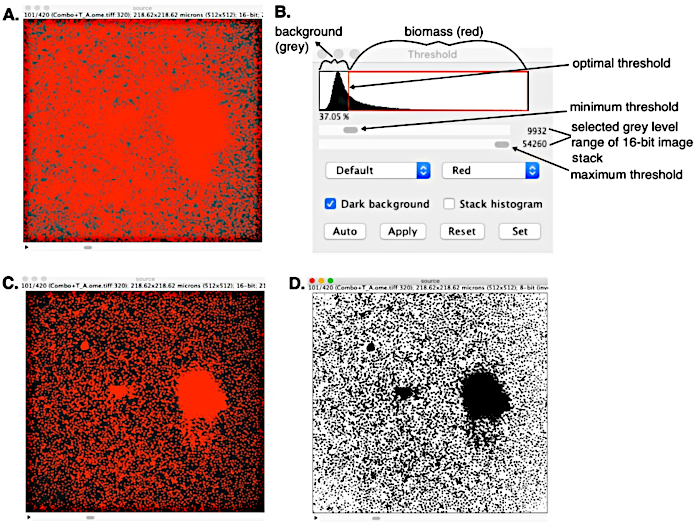
Figure 3: Simple histogram thresholding of a 16-bit CLSM OME-TIFF in ImageJ. (A) OME-TIFF before thresholding with background noise and two distinct phases: red biomass and greyscale background. (B) Threshold window used to remove background noise and set optimal threshold by converting lower intensity pixels (values less than the minimum set threshold) into black background. Maximum threshold value set to establish gray level range to be converted to red biomass. (C) Quality segmented z-stack image generated, and (D) converted stack to a binary black-and-white OME-TIFF that is readable to COMSTAT algorithms. Please click here to view a larger version of this figure.
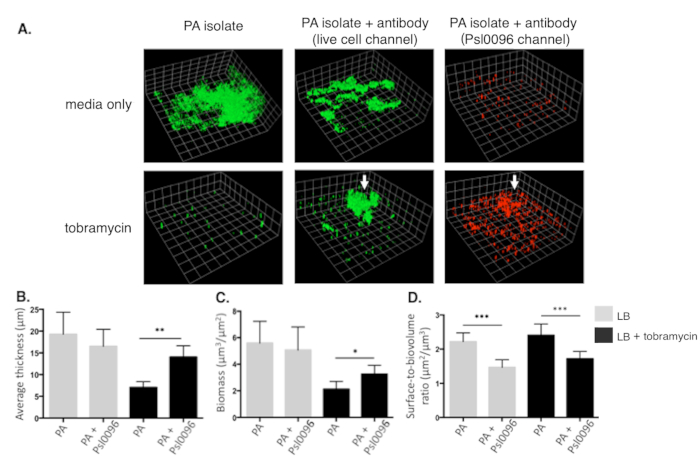
Figure 4: Representative CLSM images and COMSTAT comparison of fluorescent labelled CF Pseudomonas aeruginosa biofilms under varying antimicrobial conditions. (A) Three-dimensional images of 48 h biofilms grown in chambered coverglass. Live-cell (green) channel of biofilm grown in LB media alone (top left) and with 1000 μg/mL tobramycin in LB (bottom left), representing antibiotic effect on control wells. Live-cell channel of biofilm exposed to anti-Psl mAb, Psl0096, in the absence (top middle) and presence (bottom middle) of tobramycin, showing the formation of aggregates and tobramycin resistance. Corresponding Psl0096 (red) channel of identical biofilm is shown in the absence (top right) and presence (bottom right) of tobramycin. White arrows indicate high Psl-antibody binding localized in the same region of aggregates shown in the live cell channel. Quantitative comparison of COMSTAT parameters are displayed in a series of bar graphs, including (B) average thickness (entire well), (C) biomass, and (D) surface-to-biovolume ratio of the isolate. One scale unit is equivalent to 19.68 μm. Each bar represents average of multiple images (n=18) from 3 biologically independent experiments, in the absence (grey bars) and presence (black bars) of tobramycin, with standard error bars of the mean. *p <0.01 **p <0.001 ***p < 0.0001 by Mann-Whitney test. Please click here to view a larger version of this figure.
