To load islets into microplate, 15 islets should be aspirated in 15 µL of media, as shown in Figure 1A. Islets will naturally settle toward the bottom of the pipet tip within a few seconds. Then, the pipet tip is lowered to the bottom of the well. The tip is very slightly lifted, and a small volume (about 5 µL) is pipetted out along with the islets. This technique results in consistent placement of islet at the bottom of the microplate well (Figure 1B) allowing for accurate oxygen consumption measurements.
Figure 2 shows representative results by individual well for oxygen consumption throughout the assay. This particular experiment demonstrates what can happen when wells are loaded poorly, with many islets stuck up on the sides of the well rather than at the bottom of the well. This can be caused by pipetting too much media into the well after the islets have already been pipetted out of the tip. The flow of additional media pushes the islets out of the bottom of the well. When this happens, the baseline level of oxygen consumption will be very low, and this well will show little to no response to FCCP. The bolded line in Figure 2 demonstrates this phenomenon.
Figure 3 shows a different experiment in which most wells did show significant baseline respiration and response to drugs. However, two wells (with bolded lines) showed no response to rotenone/AA. This suggests that the drug was not properly released into the well. In this case, as with cases of no basal oxygen consumption, these wells can be excluded from further analysis.
Figure 4 shows the results of a successful assay. Here we show an example of summary data from a separate experiment in which wells were properly loaded with islets and correctly injected with drugs. ATP-dependent mitochondrial oxygen consumption was effectively inhibited by oligomycin, maximal respiration-well above basal levels-was induced by FCCP, and mitochondrial respiration was completely shut down-below oligomycin levels-by inhibition of the electron transport chain with rotenone/AA.
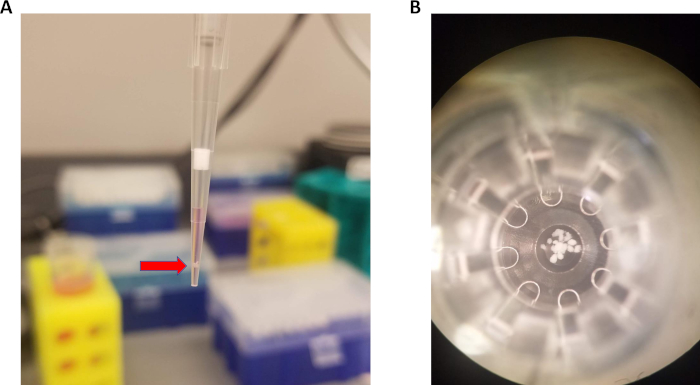
Figure 1: Pipetting technique for loading islets into microplate. (A) Approximately 15 islets (red arrow) pipetted with 15 µL of media. (B) Islets centered at the bottom of the spheroid microplate. Please click here to view a larger version of this figure.
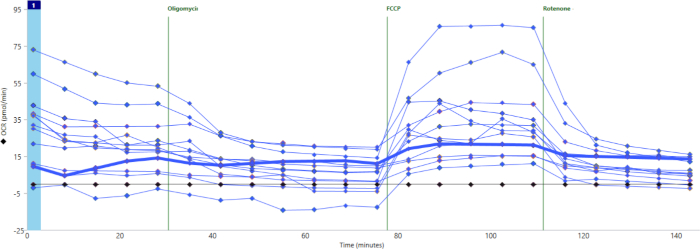
Figure 2: Well-to-well variability due to difference in islet loading. Wells loaded improperly (bold blue line) show little to no basal oxygen consumption and minimal response to cell stress test drugs. Please click here to view a larger version of this figure.
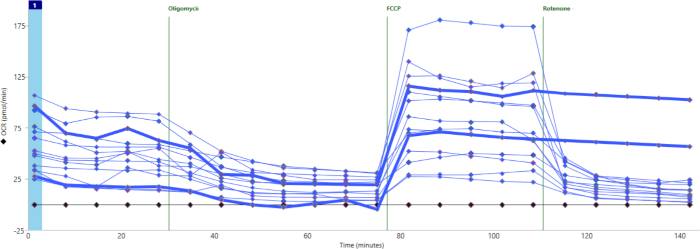
Figure 3: Failure of drug injection. Two wells (bold blue lines) were not injected with rotenone/AA and show no significant decrease in oxygen consumption. Please click here to view a larger version of this figure.
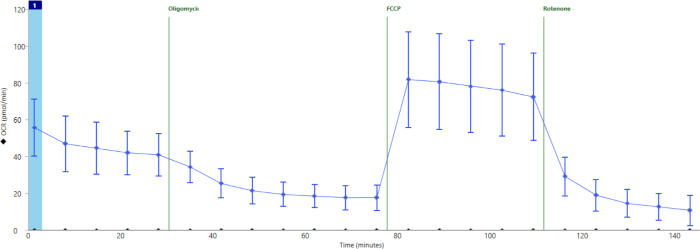
Figure 4: Averaged data across all wells in an experiment after exclusion of poorly loaded or injected wells. Average oxygen consumption throughout the cell stress test assay in a separate experiment including 16 technical replicates. Error bars show standard deviation for n = 16 wells. Please click here to view a larger version of this figure.
