The following protocol can be adapted for recombinant proteins using various affinity tags expressed in different strains of competent cells. Depending on the protein being expressed, the culturing and overnight expression conditions may require optimization of the OD600 at expression induction, expression time, expression temperature, and IPTG concentration. An overview of the protocol is illustrated in Figure 1. The details of the reagents and the equipment used in this study are listed in the Table of Materials.
1. Transformation of competent Rosetta 2 E. coli cells with the recombinant GST-USP27X expression plasmid
NOTE: To maintain the sterility of the bacterial culture, perform steps where media containers are open under a Bunsen burner flame. To allow optimal oxygen transfer, perform bacterial culture shaking in a benchtop temperature-controlled shaker with an orbit of 19 mm to 50 mm and a speed of 200 rpm29.
- Thaw 20 µL of chemically competent Rosetta 2 E. coli cells on ice immediately before use. Add 1-10 ng of the pGEX6P1-USP27X expression plasmid (encoding for N-terminal Glutathione-S-transferase (GST)30 -tagged USP27X and containing Ampicillin resistance) and incubate 5 min on ice.
- Heat shock for 30 s at 42 °C in a dry bath. Incubate on ice for 2 min.
- Add 80 µL of room temperature (RT) SOC medium. Incubate for 60 min at 37 °C with 200 rpm rotation in a 19 mm orbit benchtop temperature-controlled shaker.
- Plate 50 µL of culture on an LB agar plate supplemented with 25 µg/L Chloramphenicol and 50 µg/L Ampicillin. Incubate for 20 h at 37 °C lid-side down in a temperature-controlled incubator.
2. Overnight bacterial expression of recombinant protein from expression plasmid
NOTES: To maintain the sterility of the bacterial culture, perform steps where media containers are open under a Bunsen burner flame. To allow optimal oxygen transfer, perform bacterial culture shaking in a benchtop temperature-controlled shaker with an orbit of 19 mm to 50 mm and a speed of 200 rpm29. Measure culture OD600 using a spectrophotometer.
- Prepare 1 L of Terrific Broth (TB) medium by adding 47.6 g of TB Powder to 1 L of ultrapure water with 0.4% glycerol in a 2 L baffled culture flask. Autoclave medium for 30 min and cool to RT.
- Pick a single colony of transformed bacteria and add to 10 mL of sterile LB medium supplemented with 25 µg/L Chloramphenicol and 50 µg/L Ampicillin. Incubate for 20 h at 37 °C with 200 rpm rotation in a 19 mm orbit benchtop temperature-controlled shaker.
- Add 10 mL overnight culture to 1 L of TB medium supplemented with 25 µg/L Chloramphenicol and 50 µg/L Ampicillin (1:100 ratio of starting culture to expression culture). Incubate at 37 °C with 200 rpm rotation in a 19 mm orbit benchtop temperature-controlled shaker until the culture OD600 is between 0.5-0.6.
- Add 50 µM of IPTG to the culture to induce expression, cool to 16 °C, and incubate for 20 h at 16 °C with 200 rpm rotation.
- Pellet cells from expression culture by centrifugation for 20 min at ≥3,000 x g and 4 °C. Store the pellet at -80 °C for at least 1 h.
NOTE: At this point, the experiment can be paused and restarted later (preferably the same week). The pellet can be stored for the long term at -80 °C.
3. Protein purification by gravity-flow affinity column
NOTE: The resin, binding, wash, elution, and storage buffers appropriate for each purification will depend on the recombinant protein being purified. Collect samples from the cell pellet, supernatant, flow-through, wash fractions, and elution fractions in SDS-PAGE buffer. Perform SDS-PAGE and Coomassie staining for the samples to evaluate the success of the purification. Perform purification at 4 °C and handle fractions on ice. Cleavage of protein tags can be performed on or off the column using the appropriate protease to target the relevant protease-specific cleavage site.
- Secure the empty gravity flow column in the retort stand and fill it with glutathione agarose resin. Use 2-3 mL of resin to purify a cell pellet collected from 1 L of expression culture.
- Wash the column with one resin-bed volume of 20% ethanol. Wash column 3 times with one resin-bed volume of MS500 buffer (20 mM of Tris pH 7.5, 500 mM of NaCl, 0.5 mM of TCEP). Stop the column so the resin remains covered with buffer to prevent drying while preparing the cell pellet.
- Thaw the cell pellet at 4 °C. Add 30 mL of MS500 buffer (20 mM of Tris pH 7.5, 500 mM of NaCl, 0.5 mM of TCEP, 60 mg of lysozyme, and one protease inhibitor tablet) to the thawed pellet. Lyse for 30 min at 4 °C with gentle end-over-end rotation.
- Sonicate lysed cells in either a 50 mL centrifuge tube or a metal beaker on ice until the lysate flows freely when dispensed from a pipette tip. Centrifuge for 30 min at 4 °C and ≥20,000 x g to clear the supernatant.
NOTE: Determine sonicator settings empirically. Set the sonicator such that 120 s total time is enough to reduce the viscous lysate to a free-flowing and transparent liquid. - Decant the supernatant into a beaker and load cleared lysate onto the column. Run lysate through the column by gravity flow while collecting the flow through. Load the column with the collected flow through and run it through the column.
- Wash the column with at least two resin-bed volumes of MS500 wash buffer (20 mM of Tris pH 7.5, 500 mM of NaCl, 0.5 mM of TCEP) 5 times. Collect the wash flow through in 5 mL fractions. Add 1 µL of each fraction to 100 µL of Bradford reagent to visually check for protein presence. Wash until unbound protein is no longer present in the last wash, adding additional wash steps if necessary.
- Recover protein by running MS500 elution buffer (MS500 buffer supplemented with 10 mM of glutathione and 10 mM of NaOH) through the column, collecting 5 mL elution fractions. Check for protein presence by adding 1 µL of eluate to 100 µL of Bradford reagent. Stop collecting fractions when the Bradford reagent no longer indicates proteins are present.
- Perform buffer exchange by precipitation and centrifugation. Precipitate protein by adding 2 volumes of 4 M of ammonium sulfate to the eluate, gently inverting until cloudy, and then centrifuging for 30 min at 4 °C and ≥20,000 x g and again for another 5 min. Remove the supernatant after each centrifugation without disturbing the protein pellet.
- Re-dissolve and store protein in MS500 supplemented with 25% glycerol. Store protein pellets or protein in storage buffer at -80 °C.
4. In vitro ubiquitin chain cleavage assay
NOTE: Select ubiquitin chain length and linkage types based on DUB specificity described in previous reports31 or determined empirically. If necessary, this protocol could be used to test the activity of the wild-type DUB of interest on a panel of commercially available ubiquitin chains of defined length and linkage type. A di-ubiquitin chain amount of 375-750 ng and a DUB concentration of 1-2 µM can be used as starting points for the assay27.
- Prepare 10x DUB activation buffer (500 mM of Tris-HCl pH 7.5, 500 mM of NaCl, and 100 mM of TCEP).
- For each time point for each DUB, prepare 10 µL total of 2 µM of purified GST-USP27X in 1x DUB activation buffer (DUB mix). Prepare master mixes and split them into time points.
- Incubate the DUB mix for 10 min at RT.
- Add 7 µL of SDS-PAGE loading buffer to time 0 before adding ubiquitin chains to prevent the deubiquitylation reaction from starting.
- To each time point for each DUB, add 375 ng of K-63 di-ubiquitin chains diluted in 10 µL 1x DUB activation buffer. The total volume is 20 µL per reaction.
- Incubate the tubes at 30 °C, stopping each time point with 7 µL of SDS-PAGE loading buffer.
- Perform SDS-PAGE with a 4%-12% gradient gel and immunoblot7,32 for ubiquitin and USP27X to analyze the change in mono-ubiquitin presence across selected time points.
Figure 1: Schematic of the study design. (A) Transformation of competent E. coli cells with recombinant protein expression plasmid. (B) Overnight bacterial expression of recombinant deubiquitylase protein. (C) Protein purification of recombinant deubiquitylase using a gravity-flow affinity column. (D) In vitro ubiquitin chain cleavage assay to evaluate deubiquitylating activity. Please click here to view a larger version of this figure.
To determine the impact of XLID105-associated variants on USP27X catalytic activity, GST-tagged wild-type USP27X and the XLID105-associated variant F313V, Y381H, and S404N USP27X proteins were purified from bacteria. These variants are located within the USP catalytic domain of USP27X (Figure 2A). Because USP27X was previously reported to cleave K63 ubiquitin chains31, wild-type USP27X and the XLID105-associated variant F313V, Y381H, and S404N USP27X proteins were incubated with K63-linked di-ubiquitin chains for 1 h. These samples were separated via SDS-PAGE32, and ubiquitin and USP27X proteins were analyzed via immunoblotting. Wild-type USP27X induces di-ubiquitin cleavage, generating mono-ubiquitin (Figure 2B and Supplementary Figure 1). XLID105-associated F313V, Y381H, and S404N USP27X variant proteins did not cleave these chains. Because F313V, Y381H, and S404N variants disrupt USP27X catalytic activity, USP27X functional disruption appears to be the major mechanism underlying XLID1057. Additional quantification and complementary experiments were previously reported7.
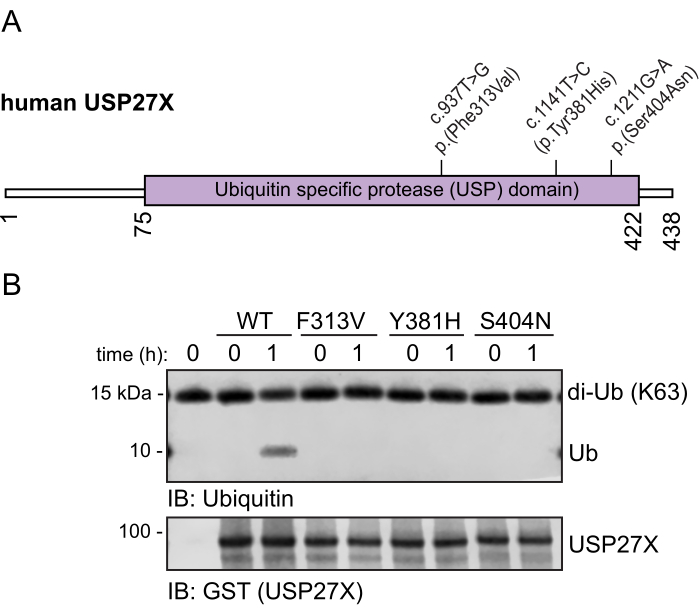
Figure 2: Impact of XLID105 variants on USP27X deubiquitylating activity. (A) Diagram of the human USP27X protein structure, showing residue numbers and the location of the XLID105 variants evaluated in the USP domain (purple). (B) GST-tagged wild-type USP27X and XLID105 variants (F313V, Y381H, and S404N) were incubated with K63 di-ubiquitin chains for the indicated times. Immunoblot analysis was performed using anti-GST (USP27X) and ubiquitin antibodies. Please click here to view a larger version of this figure.
Supplementary Figure 1: Uncropped blots of Figure 2B. Please click here to download this File.
