The workflow of the protocol is summarized in Figure 4. For this 3D organoid culture model, we adopted a thrombin and fibrinogen hydrogel that had previously been used by our team to maintain primary mouse mTECs for a couple of days, thanks to the physical and mechanical cues it provided34. After polymerization, the gel should display a loose, sponge-like mesh structure (Figure 5).
After the initial seeding and attachment phase, the organoids progressively grew and developed both at the surface and within the uppermost layers of the gel. Depending on the gel properties, the seeding conditions, and the number of organoids seeded on the gel, the organoids formed spheric to oblong structures (Figure 6) and occasionally merged to form larger structures. Two particular sub-levels of the organization were observed within the organoids after the first week of culture: first, we observed long, cell-surface projection-like structures formed by large cells irradiating from the organoids and colonizing the hydrogel in all directions (Figure 6 and Figure 7). Second, we observed cluster-like structures formed by smaller cells concentrating around those cell projections. Although we were not able to isolate both cell types to confirm the study hypothesis, this phenomenon is reminiscent of 3D arrangements found within the thymic cortex, formed by the interaction of individual cTECs with a large number of much smaller developing T cells, known as thymic nurse cell complexes11 (Figure 8).
At several time points during the organoid culture phase, we evaluated the cellular composition of the thymic organoids by flow cytometry and identified several key compartments: TEC (characterized as EPCAM+ CD45-), thymocytes (EPCAM- CD45+ CD3+) (Figure 9), as well as an EPCAM- CD45+ CD3- compartment comprising thymic hematopoietic non-thymocyte subsets. Further details can be found in Provin et al.23.
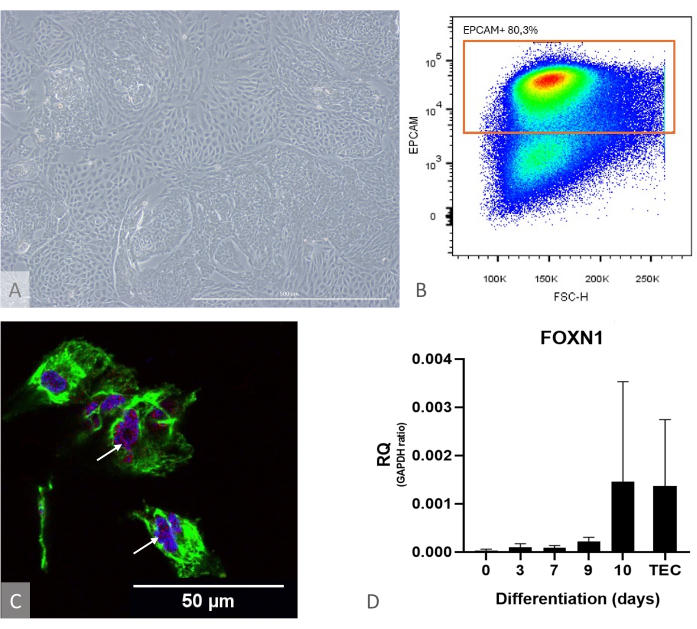
Figure 1: iPSC to TEP differentiation characterization. (A) Example of iPSC to TEP differentiation at D13, inverted phase contrast microscope, 400x. Scale bar: 500 µm. (B) Dot plot example, the proportion of EPCAM+ cells among DAPI- cells on day 14 of differentiation, image from FlowJo 10.0.7. (C) Immunostaining against DAPI (blue), PAX9 (red), and KRT8 (green), immunofluorescence, and confocal imaging on day 16 of iPSC to TEP differentiation. White arrows point to examples of anti-PAX9 staining. Scale bar: 50 µm (D) Expression level of FOXN1 (RQ to GAPDH) during iPS to TEP differentiation. TEC: Positive control reference, primary human TECs isolated from pediatric thymus samples. Graph from Prism (GraphPad version 8.0.1). Please click here to view a larger version of this figure.
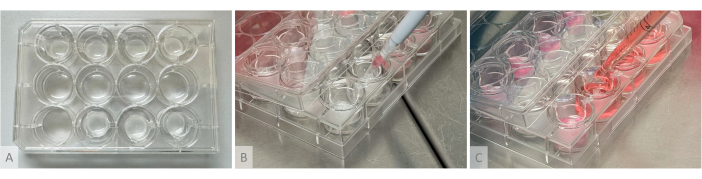
Figure 2: Experimental setup for hydrogel formation, organoid seeding, and culture medium distribution. (A) Culture plate with hydrogels casted into hanging inserts placed in the top and bottom rows. (B) Organoid seeding: the cut pipette cone containing 1 organoid is placed above the hydrogel without touching it, and the organoid is gently seeded at the surface of the gel. (C) Culture medium is deposited in the culture well by positioning the tip of the pipette between the arms of the hanging insert. Please click here to view a larger version of this figure.
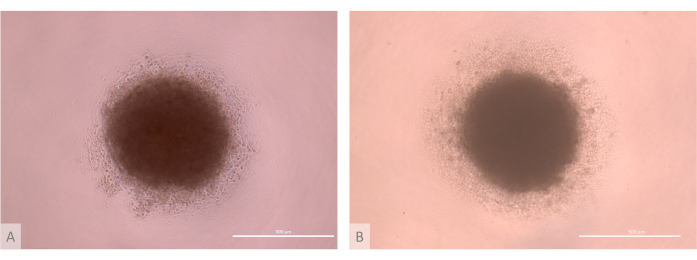
Figure 3: D0 of thymic organoid culture before seeding (days 13-15 of the full protocol). (A) organoid produced with TECs derived from the Lon71.019 iPS line. (B) Organoid produced with TECs derived from the MIPS203.003 iPS line. Inverted phase contrast microscope, 1000x. Scale bars: 500 µm. Please click here to view a larger version of this figure.
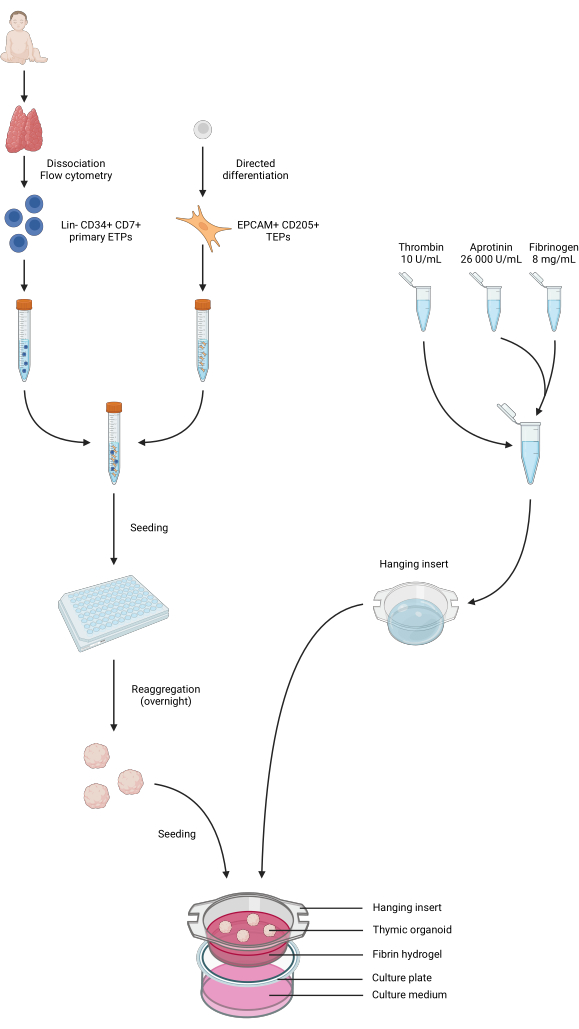
Figure 4: Summarized representation of all steps of the protocol. Pediatric thymus samples were collected and dissociated, and primary Lin- CD34+ CD7+ ETPs were sorted by flow cytometry. Differentiation of iPS cells was conducted towards a TEP identity. ETPs and iPS-derived TEPs were pooled and seeded in low-binding 96-well plates and aggregated into thymic organoids overnight. Fibrin hydrogels were prepared from aprotinin, fibrinogen, and thrombin and casted into hanging inserts. After polymerization, the organoids were seeded atop the hydrogels, and the phase 1 culture medium was added to the wells. The organoids were kept in culture for up to 6 weeks. Created in BioRender, publication license AG26EFCZOM. Please click here to view a larger version of this figure.
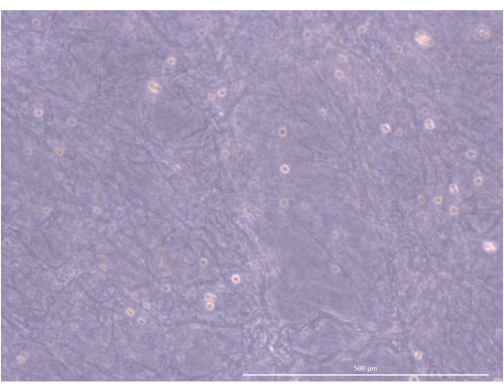
Figure 5: Organization and structure of the hydrogel. Inverted phase contrast microscope, 1000x. Scale bar: 500 µm. Please click here to view a larger version of this figure.
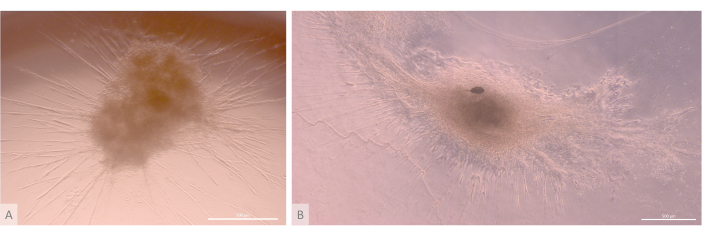
Figure 6: Mature organoids and three-dimensional structure. (A) Thymic organoid on day 24 of 3D culture, MIPS203.003 iPS line. (B) Composite image of a thymic organoid on day 32 of 3D culture, Lon71.019 iPS line. Inverted phase contrast microscope. Scale bars: 500 µm. Please click here to view a larger version of this figure.
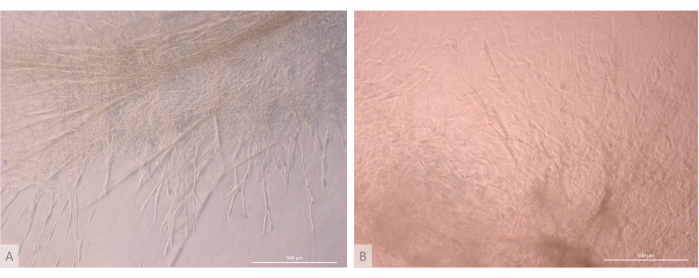
Figure 7: Structure detail of thymic organoids. (A) Thymic organoid on day 32 of 3D culture, L71.019 iPS line. (B) Thymic organoid on day 27 of 3D culture, L80.002 iPS line. Inverted phase contrast microscope, 400x. Scale bars: 500 µm. Please click here to view a larger version of this figure.
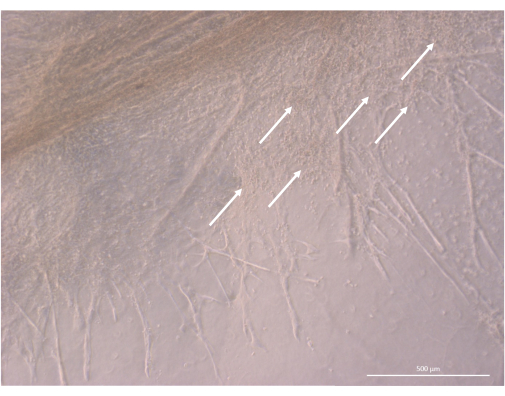
Figure 8: Structure detail of a thymic organoid on day 32 of 3D culture. White arrows point to clusters of small thymocytes proliferating in close proximity to TEC cells. Inverted phase contrast microscope, 400x. Scale bar: 500 µm. Please click here to view a larger version of this figure.
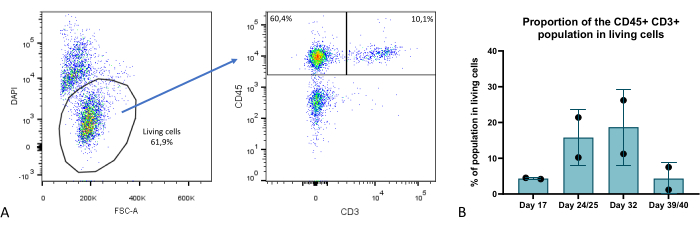
Figure 9: Proportion of the T cell compartment within thymic organoids. (A) Dot plot example, proportion of CD45+ CD3+ cells within the living (DAPI-) cells in thymic organoids on day 35 of 3D culture, image from FlowJo 10.0.7. The CD45+ CD3- fraction comprises hematopoietic non-thymocyte cells. (B) The proportion of CD45+ CD3+ cells within living cells in thymic organoids on days 17, 24/25, 32, and 39/40 of 3D culture, n=2 in technical duplicate or triplicate, graph from Prism (GraphPad version 8.0.1). Please click here to view a larger version of this figure.
| Unit | Phase 1 medium Day 14 up to Day 18 | Phase 2 medium Day 19 onward | |
| Base | XVIVO10 | XVIVO10 | |
| BMP4 | ng/mL | 50 | |
| FGF8 | ng/mL | 10 | |
| FGF10 | ng/mL | 10 | |
| IGF1 | ng/mL | 10 | |
| EGF | ng/mL | 10 | |
| RANK L | ng/mL | 50 | 50 |
| IL7 | ng/mL | 5 | 5 |
| FLT3 L | ng/mL | 5 | 5 |
| SCF | ng/mL | 10 | 10 |
| Glutamax | ng/mL | 1% | 1% |
Table 1: Supplements and their respective concentrations.
| Aprotinin (µL) | Thrombin (µL) | Fibrinogen (µL) | Phase 1 medium | Organoids (unit) | |
| 24 well plate | 5 | 75 | 75 | 1 | 3 to 5 |
| 12 well plate | 9.2 | 138.2 | 138.2 | 1.8 | 5 |
| 6 well plate | 16 | 240.8 | 240.8 | 3.2 | 8 to 9 |
Table 2: Required ratios of components for preparing hydrogels and seeding organoids in 6-, 12- and 24-well plates.
