Protocol development for vesicle production
Figure 1 outlines the two different vesicle preparation methods. The THF swelling method on the left side is composed of three successive steps and results in different supramolecular assemblies of the ELP depending on the temperature. In Figure 1A epifluorescence microscopy images show vesicles assembled from BDP-R20F20 and fibrillary structures assembled from BDP-R40F20. The BuOH method, illustrated on the right side exclusively leads to the formation of ELP vesicles, yielding about two orders of magnitude more vesicles compared to the THF swelling method. The schematic illustration shows the preparation process of BuOH vesicles. For vesicle preparation in Figure 1B BDP-R40F20 was mixed with 10-15% (v/v) BuOH and vesicles were prepared via extrusion of the mixture.
Guiding supramolecular self-assembly into different structures
Figure 2 shows a schematic illustration and epifluorescence images of different supramolecular structures assembled from BDP-R40F20 via the THF swelling protocol. In this case lyophilized BDP-R40F20 was used for the different assembly protocols. The pH of the buffer and the temperature of the assembly process were adjusted to form either coacervates, fibrils or vesicles. The coacervates depicted in Figure 2A are 1–2 µm in diameter and were assembled from BDP-R40F20 at 20 °C and pH 13. Adjustment of the assembly temperature to 90 °C results in the formation of nanofiber bundles (Figure 2B) at pH 4–13 tested with BDP-R40F20. Stable vesicles could be assembled from the ELP at a temperature of 50 °C and pH 7 (Figure 2C). Small mistakes at one of the crucial steps in the assembly protocol can lead to the formation of aggregates depicted in Figure 2D.
Encapsulation of different cargo
Figure 3 shows the encapsulation of different cargo into the vesicle lumen of vesicles assembled from F20R20-mEGFP via the BuOH extrusion method. For the encapsulation of the positively charged dye Atto Rho13 in Figure 3A, the dye was mixed with the aqueous ELP solution before addition of (15% v/v) BuOH and syringe extrusion of the mixture. The confocal microscopy images show the vesicles formed from F20R20-mEGFP in the green channel, the red dye AttoRho13 in the red channel and the resulting merged channel shows the successful encapsulation inside the vesicle lumen.
The polysaccharide Dextran Red 3000 was successfully encapsulated using the BuOH extrusion method as described above. Images recorded in green channel depict the vesicles formed from F20R20-mEGFP while red channel shows the polysaccharide cargo. Merged green and red image in Figure 3B confirm the successful Dextran Red 3000 encapsulation in to the vesicle lumen.
Membrane component compatibility and phase separation of mixed BuOH vesicles before/after extrusion
Figure 4 shows the phase separation and fusion behavior of ELP amphiphiles upon mixing of single PMBC building blocks versus assembled PMBC populations. Mixing amphiphilic ELP building blocks (F20R20-mEGFP and F20R20-mCherry) prior to PMBC assembly leads to homogenously distributed molecules within the assembled PMBC membrane. The homogenous distribution of the fluorophores and associated ELP amphiphiles is evident upon merging the red and green channel of the respective fluorescence images. By mixing vesicle populations assembled from either F20R20-mEGFP or F20R20-mCherry clearly visible membrane patches of red or green fluorescence are evident immediately after mixing. This indicates that PMBC fusion events of differently labeled PMBCs occur and that these fusing membranes and their constituents stay phase separated for at least 20 min. A similar phase behavior is known from lipid rafts, within lipid membranes23.
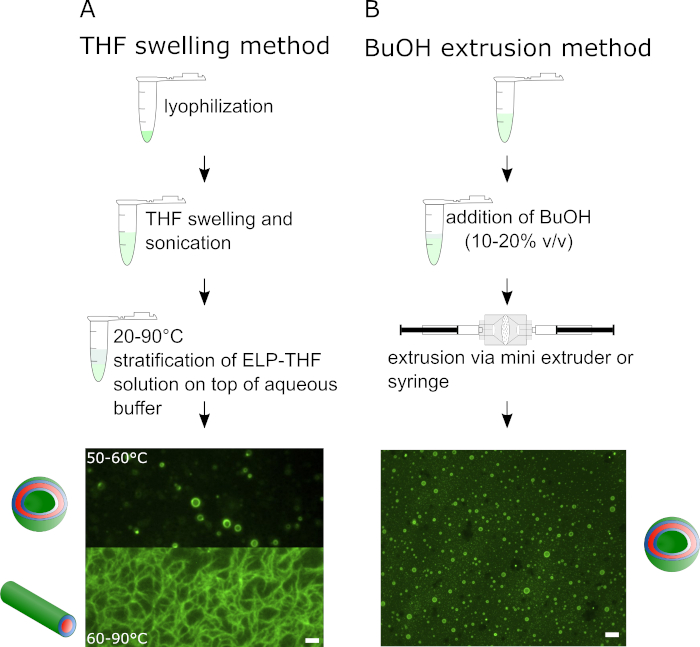
Figure 1: Illustration of the THF swelling method and the BuOH extrusion method for the guided self-assembly of amphiphilic ELPs into supramolecular structures such as vesicles or fibers. Schematic workflow and representative epifluorescence images of (A) the THF swelling method with BDP-R20F20 and BDP-R40F20 resulting in different supramolecular structures depending on temperature and pH and (B) the BuOH extrusion method exclusively yielding vesicles from BDP-R40F20 (scale bar 2 µm). This figure has been modified from Schreiber et al. 20198. Please click here to view a larger version of this figure.
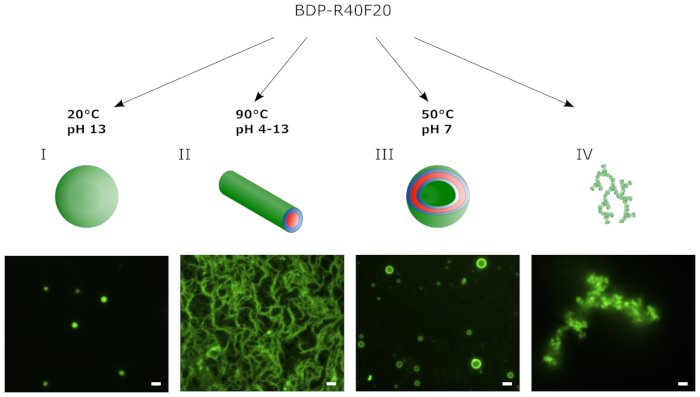
Figure 2: By applying the THF swelling method BDP-R40F20 self-assembles into different supramolecular structures. The environmental conditions applied during the assembly protocol (e.g. temperature or pH) determine the predominate supramolecular structure formed. Representative supramolecular structures at the respective conditions during the assembly were monitored via epifluorescence microscopy and range from (A) coacervates and (B) fibrils to (C) stable vesicles. (D) Failure in the assembly of defined structures during the THF swelling protocol leads to the formation of unspecific aggregates (scale bar 2 µm). This figure has been modified from8. Please click here to view a larger version of this figure.

Figure 3: Different cargos can be encapsulated within ELP vesicles using the BuOH extrusion method. (A) shows representative confocal images of F20R20-mEGFP vesicles with encapsulated positively charged dye AttoRho13 and (B) the encapsulation of the polysaccharide dextran red (scale bar 5 µm). Please click here to view a larger version of this figure.

Figure 4: Membrane component compatibility and fusion behavior of vesicle membranes assembled from F20R20 via BuOH extrusion method. (A) Mixing of fluorescent F20R20-mEGFP and F20R20-mCherry protein solution prior to syringe-extrusion leads to PMBC membranes with homogenously distributed amphiphilic proteins visible in green channel (left image), red channel (middle image), and merged channel (right image). (B) PMBCs assembled from either F20R20-mEGFP or F20R20-mCherry and mixed subsequently via syringe extrusion lead to separated ELP amphiphile patches within the PMBC membranes. The separated ELP amphiphiles within the membrane are visible after PMBC fusion for at least 20 min in green channel (left image), red channel (middle image), and the merged channel (right image). Scale bars correspond to 5 µm. This figure has been modified from Schreiber et al. 20198. Please click here to view a larger version of this figure.
